Bài giảng Thực vật thủy sinh | Đại học Quảng Bình
Bài giảng Thực vật thủy sinh | Đại học Quảng Bình. Tài liệu được biên soạn dưới dạng file PDF gồm 60 trang, giúp bạn tham khảo, ôn tập và đạt kết quả cao trong kì thi sắp tới. Mời bạn đọc đón xem!
Môn: Thực vật thủy sinh 1 tài liệu
Trường: Trường Đại học Quảng Bình 26 tài liệu
Tác giả:


























































Preview text:
TRƯỜNG ĐẠI HỌC QUẢNG BÌNH
KHOA NÔNG – LÂM – NGƯ BÀI GIẢNG
(Lưu hành nội bộ)
THỰC VẬT THỦY SINH
(Dành cho ngành Nuôi trồng thủy sản)
Biên soạn: Phan Thị Mỹ Hạnh Năm 2012 1
CHƯƠNG 1. MỞ ĐẦU
I. ĐỐI TƯỢNG, NHIỆM VỤ CỦA MÔN HỌC
Thực vật thủy sinh là một môn học chuyên nghiên cứu về các loài thực vật
sống trong môi trường nước, sự đa dạng của chúng cũng như mối quan hệ giữa
các loài với nhau và với môi trường sống. Bài giảng này chỉ giới thiệu những đối
tượng phổ biến và có vai trò quan trọng trong nghề Nuôi trồng thủy sản.
Nhiệm vụ của môn học là trang bị cho sinh viên những kiến thức cơ bản
về đặc điểm sinh học của các nhóm thực vật thủy sinh và vai trò của chúng đối
với nghề Nuôi trồng thủy sản.
II. PHƯƠNG PHÁP NGHIÊN CỨU PHÂN LOẠI THỰC VẬT 1. Phương pháp sinh học
- Phương pháp nghiên cứu hình thái (hay phương pháp so sánh hình thái)
Đây là phương pháp nghiên cứu hình dạng bên ngoài của thực vật, bao
gồm cơ quan dinh dưỡng và cơ quan sinh sản. Phương pháp này được sử dụng
phổ biến, đơn giản, ít tốn kém nhưng số liệu thu được khó chính xác vì những
các cơ quan của cơ thể luôn biến đổi tùy theo điều kiện môi trường bên ngoài. Ví
dụ sự tiêu giảm sừng ở tảo lục Scenedesmus trong môi trường pH thấp và hàm
lượng đạm giảm. Do vậy phương pháp này cần có sự hỗ trợ của những phương
pháp khác để đảm bảo đem lại kết quả chính xác nhất.
- Phương pháp giải phẫu
- Phương pháp bào tử phấn hoa: nghiên cứu về bào tử và hạt phấn.
- Phương pháp tế bào học: nghiên cứu về số lượng, hình thái tế bào và cấu
tạo bộ nhiễm sắc thể.
- Phương pháp nuôi cấy: sử dụng rộng rãi đối với tảo và nấm. Dựa vào đặc
tính của mỗi loài chỉ có thể sinh trưởng trên những môi trường chọn lọc.
2. Phương pháp địa cư
- Phương pháp địa lý thực vật: nghiên cứu khu phân bố của thực vật. Mỗi
loài có một phạm vi phân bố riêng. Vùng phân bố ảnh hưởng đến tính thích nghi
và lịch sử phát triển của mỗi loài thực vật.
- Phương pháp sinh thái học: nghiên cứu sự thay đổi, biến dị của loài do
ảnh hưởng của các điều kiện môi trường sống.
3. Những quy định quốc tế về hệ thống phân loại thực vật
Taxon và bậc phân loại
Taxon là một nhóm cá thể được coi như đơn vị hình thức ở bất kỳ mức độ
nào của thang chia bậc hay nói cách khác taxon là một nhóm sinh vật có thật
được chấp nhận làm đơn vị phân loại ở bất kỳ mức độ nào.
Bậc phân loại dùng để chỉ mức độ của taxon, là một tập hợp mà các thành
viên của nó là các taxon ở một mức nhất định trong thang chia bậc đó.
Các bậc phân loại cơ bản của giới thực vật: Giới: Regnum Ngành: Divisio Lớp: Classis Bộ: Ordo Họ: Familia Chi: Genus 2 Loài: Species Thứ: Varietas Dạng: Form
Trong các bậc phân loại nói trên, loài được xem là bậc cơ sở vì duy nhất
chỉ có bậc này quan hệ tương ứng với các chủng quần có thật trong tự nhiên. Các
bậc khác chỉ mang ý nghĩa độ xa gần trong quan hệ họ hàng của lịch sử phát sinh thế giới thực vật.
III. NHỮNG THÀNH TỰU NGHIÊN CỨU VỀ THỰC VẬT THỦY SINH Ở VIỆT NAM
Việc nghiên cứu thực vật thủy sinh đã được thực hiện tương đối sớm ở
Việt Nam. Nhìn chung, trước Cách mạng tháng Tám, việc nghiên cứu thực vật ở
nước chỉ mang tính chất điều tra khu hệ, phân loại khái quát. Các công trình
nghiên cứu đáng chú ý trong thời gian này là:
- Công trình điều tra vịnh Nha Trang của Rose (1926) đã phân loại rất
khái quát, xác định được 42 loài thực vật nổi.
- Fere (1933) xác định được 43 loài Flagelles ở nước ngọt thuộc vườn Bách thảo Sài Gòn.
Sau khi giải phóng miền Nam, chính phủ nước ta đã coi trọng việc thăm
dò nghiên cứu nguồn lợi thủy sản nước ta. Có nhiều tổ chức, cơ sở nghiên cứu
như Viện nghiên cứu Hải sản, Trường đại học Tổng hợp Hà Nội, Trường đại học
Thủy sản Nha Trang... tiến hành công tác nghiên cứu. Lúc này, các công trình
không chỉ dừng lại ở điều tra cơ bản, phân loại đơn thuần mà đã có những
nghiên cứu về sinh lý, sinh thái và tiến hành nuôi một số loài tảo có giá trị kinh
tế như Chlorella, Spirulina, Skeletonema... Những công trình nổi bật ở giai đoạn này là:
- Công trình điều tra tổng hợp vịnh Bắc bộ năm 1959-1963 của đoàn điều
tra Việt – Xô, Việt – Trung.
- Công trình nghiên cứu của Trương Ngọc An và Hàn Ngọc Lương (1970-
1971) ở cửa sông Ninh Cơ (Nam Hà) đã giám định được 115 loài thực vật nổi.
- Công trình nghiên cứu phiêu sinh vật trong vịnh Nha Trang của Hoàng
Quốc Trương (1962-1963) giám định được 112 loài tảo Silic, 92 loài tảo Giáp.
- Vũ Trung Tạng và Hoàng Thị Sy (1976) xác định được 86 loài thực vật
nước ở đầm phá Nam sông Hương.
- Vũ Thị Tám và Nguyễn Trọng Nho (1978-1980) xác định được 135 loài
thực vật nổi ở đầm Thị Nại (Bình Định), 116 loài thực vật nổi ở đầm Nha Phu (Khánh Hòa).
- Trương Ngọc An (1993) phân loại ngành tảo Silic ở Việt Nam.
IV. VAI TRÒ CỦA THỰC VẬT THỦY SINH
1. Đối với tự nhiên
Thực vật thủy sinh là bộ phận cấu thành sinh giới. Thực vật ở nước cũng
giống như thực vật trên cạn, chúng có khả năng quang hợp, tổng hợp nên các
chất hữu cơ từ các vật chất vô cơ. Biển và đại dương chiếm một phần rộng lớn (361.106 km2 6 2
) hơn rất nhiều so với diện tích lục địa (149.10 km ). Hằng năm,
các loại rong tảo tổng hợp được 13,5.1010 tấn vật chất hữu cơ, trong khi toàn bộ 3
thực vật ở lục địa chỉ tổng hợp được 1,9.1010 tấn. Như vậy có thể nói thực vật
thủy sinh là nhân tố chính hấp thụ CO2 và thải O2, điều hòa cân bằng không khí.
Các thàm thực vật phát triển thành rừng dưới đại dương hay trong hệ sinh
thái rừng ngập mặn vừa là nơi trú ẩn vừa là nơi cung cấp thức ăn cho các sinh
vật ở nước cũng như trên cạn. Các hệ sinh thái ở nước còn là kho lưu giữ đa
dạng sinh học, nuôi dưỡng các loài sinh vật quý, hiếm.
Ngoài ra, thực vật thủy sinh còn có vai trò quan trọng trong các chu trình
chu chuyển vật chất của tự nhiên (chu trình C, chu trình N...), giúp cho vật chất
được trao đổi liên tục.
2. Đối với con người
Hầu hết các loài rong tảo biển không chứa độc tố, một số giống loài có
hàm lượng dinh dưỡng cao đã được con người khai thác và sử dụng làm thức ăn
rất phổ biến ở các nước châu Á (Nhật Bản, Trung Quốc, Philippin...). Số giống
loài làm thức ăn khá nhiều: thực vật nổi có Nosto, Spirulina maxima, Chlorella
pyrenoidora... thực vật đáy có Eucheuma, Gracilaria, Porphyra, Ulva,
Enteromorpha... Với đặc tính giàu protein, vitamin (nhất là vitamin C), giàu các
nguyên tố khoáng như Iốt, Ca, Fe, Co nhưng ít chất béo nên rong biển ngày càng
được ưa chuộng, nhất là đối với những người ăn kiêng.
Rong biển còn được sử dụng để chiết xuất các loại keo rong, phục vụ cho
các ngành công nghiệp thực phẩm, đồ hộp. Keo algin và các muối alginat của
chúng được sử dụng nhiều trong khâu in vải của ngành công nghiệp dệt, trong kỹ
nghệ mỹ phẩm... một số loài rong (Cladophora) được dùng làm nguyên liệu sản
xuất giấy. Một số loại khác được dùng làm nguyên liệu sản xuất hóa chất, dược phẩm.
Nhiều loài rong có tác dụng chữa bệnh được dùng trong y học, điều chế
thuốc kháng sinh (Cyanobacteria, Chlorella chiết rút chất độc dùng làm chất
kháng sinh, kháng ký sinh trùng...).
Nói chung, thực vật thủy sinh có ảnh hưởng rất lớn đến đời sống con
người. Nếu không có thực vật thủy sinh (đặc biệt là rong tảo) sẽ không có kinh tế
biển, toàn bộ chu trình vật chất và cuộc sống trên trái đất sẽ bị hủy diệt.
3. Vai trò của thực vật thủy sinh đối với nghề Nuôi trồng thủy sản
Thực vật thủy sinh nói chung và thực vật nổi nói riêng có vai trò rất quan
trọng đối với nghề nuôi thủy sản. Thực vật nổi là nguồn thức ăn tự nhiên không
thể thay thế của nhiều đối tượng thủy sản. Nhiều loài trong ngành tảo silic, tảo
lục là thức ăn tốt cho ấu trùng của các loài cá, giáp xác, thân mềm... Có thể nói
nếu không có thực vật nổi thì không có nghề nuôi cá. Nhờ có thực vật thủy sinh
mà khâu thức nhất trong quá trình tạo ra vật chất hữu có trong thủy vực được
thực hiện. Năng suất sơ cấp của thủy vực là khâu quan trọng, quyết điịnh năng
suất của thủy vực,là cơ sở của quá trình tạo thành những chất sống ở những bậc cao hơn sau này.
Bên cạnh đó, một số thực vật ở nước còn gây tác hại đối với các đối tượng
thủy sản như tảo Silic hình thuyền Amphora sống bám trên mang tôm thịt gây
bệnh đen mang hoặc sự phát triển mạnh của tảo có thể dẫn đến hiện tượng “nở
hoa nước” làm ô nhiễm môi trường, gây độc cho đối tượng nuôi. 4
CHƯƠNG 2. GIỚI THIỆU CHUNG VỀ TẢO 1. Khái niệm
Tảo thuộc giới thực vật, bao gồm một nhóm sinh vật rất đa dạng, khó định
nghĩa chính xác. Sự phân chia ngành của chúng còn có nhiều ý kiến, 6 ngành, 12
hay 13 ngành ... (ở đây tạm chia thành 10 ngành). Đến nay còn một số tảo vẫn
chưa được biết đến một cách tỉ mỉ.
Tảo bao gồm cả thể tiền nhân và có nhân thật. Người ta cho rằng tảo là
nhóm sinh vật đầu tiên, từ đó nhóm thực vật không hoa, và cuối cùng là nhóm
thực vật có hoa xuất hiện.
Tảo có cấu tạo cơ thể dạng tản, dạng đơn độc hay tập đoàn, dạng sợi hay
mô mềm... Nhiều dạng đơn bào có thể chuyển động và có thể có mối liên quan
với protozoa. Về hình thái tảo rất đa dạng, một số lớn tảo nâu (Phaeophycota) có
thể đạt kích thước tương đương với một cây nhỏ. Tuy là những sinh vật tương
đối đơn giản nhưng ngay trong những tế bào nhỏ nhất cũng thể hiện một sự hoàn
hảo ở cấp độ tế bào.
Tóm lại, tảo là những thực vật bậc thấp, cơ thể chưa có sự phân hóa thành
thân, rễ, lá (những dấu hiệu của thực vật bậc cao) nên cơ thể chúng được gọi
chung là tản. Cơ thể chúng có chứa sắc tố quang hợp, chúng có khả năng quang
tự dưỡng sử dụng năng lượng mặt trời chuyển những chất vô cơ thành dạng
đường đơn giản. Tản có cấu trúc rất đa dạng: đơn bào, tập đoàn hay đa bào.
2. Các dạng hình thái của tản
Tản của tảo có thể có các hình dạng cấu trúc sau đây:
- Dạng đơn bào: Tế bào hình trái xoan, hình quả lê hay gần như hình cầu,
êlip... Tản có thể là đơn bào hay thành tập đoàn, cấu tạo từ một số hay nhiều tế
bào giống nhau về hình thái và chức năng.
Dạng đơn bào này có thể gặp trong cả quá trình sống của những tảo đơn
giản hay ở giai đoạn sinh sản (bào tử và giao tử) của những tảo có tổ chức cao hơn.
- Dạng hạt: tế bào không có roi, thường hình cầu, đôi khi hình khác, tế
bào đơn độc hay liên kết trong tập đoàn.
- Dạng sợi: gồm các tế bào liên kết nhau thành sợi đơn hay phân nhánh.
Các tế bào trong sợi phần lớn giống nhau hay đôi khi một số tế bào ở gốc hay ở
ngọn có hình dạng và cấu tạo khác biệt.
- Dạng bản: được hình thành từ dạng sợi trong quá trình cá thể phát sinh
của tảo. Ở đây các tế bào phân chia theo cả chiều ngang và chiều dọc, kết quả
tạo nên dạng bản có hình lá rộng hay hẹp. Nhiều tảo ở biển (như Tảo nâu, Tảo đỏ) có cấu trúc này.
- Dạng ống: thường gặp ở một số tảo mà cơ thể dinh dưỡng của chúng chỉ
là một tế bào khổng lồ có kích thước tới hàng chục centimét thậm chí hàng vài
mét, chứa nhiều nhân và không có vách ngăn thành các tế bào riêng rẽ. Dạng
ống có thể đơn hoặc phân nhánh hình cành. 5
3. Ðặc điểm cấu tạo tế bào
Tảo có cấu trúc 1 tế bào thực vật gồm màng bao bọc, bên trong là nguyên
sinh chất với nhân điển hình (chất nhân, màng nhân và hạch nhân). Màng có cấu
trúc bằng xenlulo hoặc hemixenlulo, một số tảo màng có nhiễm thêm chất
SiO2.nH2O. Cấu trúc màng là một vỏ bao, bao có thể gồm 2 hoặc nhiều tấm hợp
lại như ở Bacillariophyta (màng gồm 2 tấm) và Dinophyta (màng gồm nhiều
tấm). Đặc điểm cấu trúc vỏ là tiêu chuẩn để phân loại tảo.
Trong nguyên sinh chất chứa lục lạp (Chloroplast) gồm các thylakoit riêng
rẽ hoặc liên kết với nhau. Trên các thylakoit mang các sắc tố. màng lục lạp cũng
là đặc điểm quan trọng để phân loại các ngành tảo.
Ngoài ra, trong nguyên sinh chất còn chứa các thể ribo, các hạt cơ thể,
lipit, không bào, ở tảo mắt còn có các điểm mắt (stigura), nhờ đó tế bào di
chuyển về phía ánh sáng.
Vách tế bào của tảo phần lớn được cấu tạo bởi cellulose và pectin, một vài
loài tảo vách có thấm thêm silic (như Khuê tảo, Tảo vàng ánh) hoặc cacbonat
canxi (Tảo vòng). Mỗi tế bào có một nhân hay đôi khi nhiều nhân (ở Tảo dạng
ống). Trong chất nguyên sinh có những bản (thylakoids) chứa diệp lục và các sắc
tố khác được bao bọc lại gọi là lạp. Lạp có hình dạng khác nhau, ổn định với
từng giống riêng rẽ hay là với các nhóm phân loại lớn, có thể có dạng bản, dạng
dải xoắn, hình sao, mạng lưới, dạng đĩa, dạng hạt.
Ở một số tảo như Tảo lục, trong lạp có các thể đặc biệt gọi là hạt tạo bột
(pyrenoid), là những thể protein hình cầu hay có góc, xung quanh tập trung các
hạt tinh bột hay hidratcacbon là chất dự trữ chính của Tảo lục, ngoài ra còn có
những giọt lipid ở trong hoặc ngoài lạp (như ở các tảo khác).
Ðối với những tảo có cấu trúc đơn bào, trong tế bào của chúng chứa đầy
chất nguyên sinh và không có không bào với dịch tế bào. Nhưng với những loài
sống ở nước ngọt, trong chất nguyên sinh ở phần đầu tế bào chứa một hay một
vài không bào co bóp, có chức phận thải các sản phẩm thừa trong trao đổi chất ra
ngoài tế bào, và điều chỉnh sự thẩm thấu của tế bào.
Đa số tảo là sinh vật quang dưỡng, tất cả đều có chlorophylla và có sản
phẩm cuối cùng là carbonhydrat và protein tương tự với những thực vật bậc cao
hơn. Nhiều tảo là sinh vật thí nghiệm lí tưởng nhờ vào kích thước nhỏ của chúng
và dễ dàng thao tác trong môi trường lỏng. Chúng có thể được nghiên cứu dưới
những điều kiện được kiểm soát trong phòng thí nghiệm. 4. Sinh sản
Tảo có nhiều hình thức sinh sản khác nhau: sinh sản sinh dưỡng, sinh sản
vô tính, sinh sản hữu tính; nhiều tảo có sự xen kẽ thế hệ.
* Sinh sản sinh dưỡng: thực hiện bằng những phần riêng rẽ của cơ thể
thường không chuyên hóa về chức phận sinh sản. Ở các tảo đơn bào sinh sản
sinh dưỡng thực hiện bằng cách phân đôi tế bào; ở các tảo tập đoàn thì tách ra
thành các tập đoàn nhỏ hay hình thành tập đoàn mới ở bên trong tập đoàn (như ở
Volvox); các tảo dạng sợi sinh sản sinh dưỡng bằng cách phát triển một đoạn tản 6
được tách rời khỏi tản cũ được gọi là Tảo đoạn. Một số ít tảo có tạo thành cơ
quan chuyên hóa của sinh sản sinh dưỡng như hình thành chồi ở Tảo vòng (Chara).
* Sinh sản vô tính: thực hiện bằng sự hình thành các bào tử chuyên hóa,
có roi hoặc không có roi. Các bào tử được hình thành trong bào tử phòng (túi
bào tử). Bào tử nẩy mầm thành tản mới.
* Sinh sản hữu tính: thực hiện bằng sự kết hợp của những tế bào chuyên
hóa gọi là giao tử. Tùy theo mức độ giống nhau hay khác nhau của các giao tử
mà phân biệt 3 hình thức sinh sản hữu tính là đẳng giao, dị giao và noãn giao.
Hợp tử được hình thành sau khi kết hợp giao tử hoặc thụ tinh sẽ nẩy mầm trực
tiếp thành tản mới hoặc qua giai đoạn trung gian.
5. Môi trường phân bố
Ở đâu có nước ở đó có tảo. Tảo thường sống ở trong nước ngọt hay nước
mặn, trôi nổi tự do trong lớp nước ở trên mặt, có trong thành phần của các sinh
vật phù du (plankton), cũng có khi chúng sống bám vào đáy hay các giá thể khác
ở dưới nước hoặc nằm tự do ở dưới đáy, tham gia vào nhóm sinh vật đáy
(benthos). Nhiều tảo còn sống trên cạn (trên đất, đá, thân cây...), sống trên băng tuyết...
Như vậy, tảo có mặt ở khắp mọi nơi, xuất hiện ở mọi môi trường sống trên
trái đất, từ vùng sa mạc nóng và lạnh khắc nghiệt đến vùng đất đá băng tuyết và
mọi thuỷ vực. Chúng là những sinh vật sản xuất sơ cấp, đóng vai trò chính trong
chuỗi thức ăn ở các hệ thuỷ vực. Một số tảo là nguồn thực phẩm của con người,
và chiết xuất hoá học của tảo được sử dụng trong công nghiệp thực phẩm và
nhiều ngành công nghiệp khác. Tuy nhiên chúng là "những sinh vật phiền toái"
cho các hệ thống cung cấp nước cho đô thị và trong thuỷ vực dễ bị phú dưỡng.
Chúng là những sinh vật gây nên hiện tượng nở hoa ở nước (water bloom), đặc
biệt một số tảo còn sản sinh ra độc tố gây tác hại cho con người thông qua dây
chuyền thức ăn. Trong tương lai công dụng của tảo sẽ phát triển trong lĩnh vực
năng lượng và sản xuất thực phẩm. 6. Vai trò của tảo
Tảo là thành phần sống của hệ sinh thái, nó liên hệ với các thành phần
khác qua việc tham gia vòng tuần hoàn vật chất, là mắt xích đầu tiên của chuỗi thức ăn.
Nguồn lợi từ Tảo biển đóng vai trò quan trọng trong ngành Thủy sản,
cung cấp năng lượng sạch, xử lý môi trường, nguyên liệu cho công nghiệp, y
khoa, mỹ phẩm, dược phẩm, phân bón cho trồng trọt, nguồn thực phẩm dinh
dưỡng cao cho chăn nuôi lẫn con người.
Nguồn lợi tảo đa phần là tảo nước mặn. Đóng vai trò quan trọng việc giải
quyết những vấn đề có tính chất toàn cầu của con người: Thực phẩm, năng
lượng, bảo vệ môi trường, là nguồn cung cấp nguyên liệu cho công nghiệp, vật
liệu xây dựng, y tế, mỹ phẩm, dược phẩm những chất có hoạt tính sinh học cao.
Thức ăn cho ấu trùng thủy sản. 7
Ở Việt Nam, Nuôi trồng Tảo cũng đã có từ lâu nhưng chưa phát triển
mạnh do nhiều nguyên nhân, yếu tố quan trọng nhất là giá cả tiêu thụ, canh tranh
cũng như chất lượng sản phẩm.
Một số nước như Nhật, Anh, Pháp, Mỹ, Canađa… Sản xuất Tảo biển đạt
đến 60-120 tấn/ha/năm với các loài như Porphyra, Laminaria, Chondrus, Ulva,
Undari, Monostroma, Rhodymenia, Gelidium.
Theo thống kê của FAO, mức sản xuất và khai thác Tảo biển có kích thước
lớn trên thế giới hàng năm xấp xỉ 4 triệu tấn Rong tươi, trong đó Tảo Nâu chiếm
65%, Tảo Đỏ 33%, Tảo Lục 0,4%. Ở Việt Nam, cũng đang phát triển nuôi trồng
và được xem xét dùng làm nguồn thực phẩm phụ. 8
CHƯƠNG 3. CÁC NGÀNH TẢO
I. Ngành tảo lam - CYANOPHYTA
1. Lớp tảo lam - Cyanophyceae
Tảo lam là sinh vật cổ xưa nhất. Tảo lam dạng sợi hoá thạch có tuổi ít nhất
là 3,5 tỉ năm (Schopf 1993). Mặc dầu tế bào không có cấu trúc phức tạp so với
các tảo khác nhưng tảo lam vẫn là đại diện có vai trò quan trọng ở các hệ sinh
thái. Tảo lam là sinh vật quang hợp đầu tiên tổng hợp chất hữu cơ và cũng là tế
bào đầu tiên có hai hệ thống tiếp nhận ánh sáng và giải phóng O2. Nhiều loài tảo
lam có khả năng cố định đạm, chuyển Nitơ trong khí quyển từ thể tự do sang
dạng Nitơ sử dụng được như amonium (NH4), acid amino và một loạt hợp chất nitơ khác.
Tảo lam thuộc nhóm tiền nhân, nhân không có màng nhân, không có lưới
nội sinh chất, ty thể, thể golgi, lạp thể và không mang roi.
Thành tế bào tảo lam cấu tạo bởi chất peptidoglycan, tương tự cấu tạo
thành tế bào của vi khuẩn gram âm, bao gồm hai lớp:
- Lớp peptidoglycan cứng dính liền với màng tế bào
- Lớp lipopolysaccharide nằm ở phía ngoài.
Peptidoglycan là một cao phân tử của N-acetylglucosamine và N-
acetylmuramic acid được nối kết bởi các amino acid. Thành tế bào ở một số loài
tảo lam có vỏ bao nhầy bên ngoài. a. Sinh sản
Tảo lam không có sinh sản hữu tính, chỉ sinh sản theo hình thức dinh
dưỡng bằng phân đôi tế bào và tảo đoạn; sinh sản vô tính bằng nội và ngoại bào tử. 9
Tảo lam hình thành bào tử màng dày (akinete) còn gọi là bào tử nghỉ
(resting spore), có khả năng chống chịu với điều kiện môi trường bất lợi. Bào tử
nghỉ gặp ở các bộ Nostocales, Stigonematales.
Tảo lam còn có tế bào đặc biệt gọi là dị bào (heterocyst). Dị bào lớn hơn
tế bào dinh dưỡng và trông như một tế bào trống khi quan sát dưới kính hiển vi
quang học do thiếu các hạt dự trữ và không bào khí. Tính chất đặc trưng của dị
bào là vách dày, thường ở các cực tế bào vách dày và nhô vào phía trong được
gọi là các nốt ở cực (polar nodule). Các nốt này có các kênh rất nhỏ nối liền dị
bào với tế bào dinh dưỡng kế cận. Dị bào là tế bào chuyên hoá để cố định nitơ ở khí quyển.
Trong quá trình này, nitơ từ khí quyển sẽ được chuyển thành amonium.
Khi lượng nitơ hoà tan trong môi trường xung quanh thấp, một số tế bào dinh
dưỡng sẽ được chuyên hoá để trở thành tế bào dị hình, thành của tế bào dinh
dưỡng dày lên do được bổ sung thêm một số lớp, và giữa thành tế bào dị hình
mới được tạo lập với tế bào dinh dưỡng kế cận sẽ xuất hiện nhiều lỗ thông.
Đường từ tế bào dinh dưỡng bên cạnh được chuyển vào tế bào dị hình để dùng
cho oxy hoá của quá trình hô hấp.
Tảo lam có tế bào dị hình, thường thuộc vào các bộ như Nostocales, và
Stigonematales. Mặc dầu tế bào dị hình có chức năng cố định đạm của khí quyển
nhưng một số tảo lam không có tế bào này vẫn có khả năng cố định đạm.
b. Sự phân bố của tảo Lam
Tảo Lam phân bố rộng rãi ở cả môi trường nước ngọt và biển. Chúng sống
trôi nổi hay sống bám ở đáy thuỷ vực. Một số tảo lam sống ở môi trường cạn (vỏ
cây, đất ẩm...), một số sống cộng sinh bên trong cơ thể sinh vật khác. Nhiều tảo
lam cộng sinh đã tạo ra nguồn đạm cho vật chủ và cả chính mình. Nhờ có khả
năng cố định đạm đã giúp cho tảo lam sống thuận lợi hơn các loại tảo khác trong
các thuỷ vực có nồng độ nitơ thấp.
Tảo lam có thể sống trong môi trường có nhiệt độ cao, độ kiềm cao và có
thể chống chịu được cả một thời kỳ khô hạn. Synechococcus sống được ở nhiệt
độ môi trường lên đến 740C. Ngược lại nhiều loài tảo lam sống ở các hồ vùng
bắc cực có tuyết phủ quanh năm với độ dày hàng mét. Tảo lam cũng gặp ở các
hồ, vũng ven biển có độ mặn cao do quá trình bốc hơi nước. Một số tảo lam có
thể tiến hành quang hợp trong môi trường yếm khí tương tự như vi khuẩn. c. Phân loại
Hiện nay vẫn tồn tại hai hệ thống phân loại đối với tảo lam: hệ thống phân
loại đặt tảo lam ở nhóm thực vật truyền thống do Geitler đưa ra năm 1932 và hệ
thống phân loại xếp tảo lam ở nhóm vi khuẩn do Stainier và cộng sự đưa ra vào năm 1971. 10
Theo C. Van Den Hoek (1995) lớp tảo lam gồm 5 bộ, đó là:
Chroocoicales, Pleurocapsales, Oscillatoriales, Nostocales, Stigonematales.
- Bộ Chroococcales: Tản đơn bào, đơn độc hay tập đoàn. Tế bào tròn
không phân biệt gốc và đỉnh, không có nội và ngoại bào tử.
Tế bào đơn độc hay tập đoàn nhưng không sắp xếp thành hàng hay sợi (họ
Chroococcaceae), thường gặp như: Chroococcus, Microcystis (hình 2)
- Bộ Pleurocapsales: Tản hình sợi, có phân nhánh hoặc không, sinh sản
cách phân chia tế bào hoặc nội bào tử.
Các chi điển hình: Cyanocystis, Pleurocapsa.
- Bộ Nostocales: Tản hình sợi, có sự sinh sản bằng tảo đoạn, có tế bào dị
hình và bào tử, phân nhánh giả.
Các chi điển hình: Nostoc, Anabaena, Aphanizomenon,...
- Bộ Oscillatoriales: tản dạng sợi không có tế bào dị hình, không có bào
tử, sinh sản bằng tảo đoạn. Gồm các chi: Oscillatoria, Spirulina, Lyngbya...
- Bộ Stigonematales: tản dạng sợi, phân nhánh thật, có sinh sản bằng tảo
đoạn, thường có tế bào dị hình. Chi điển hình Stigonema.
2. Lớp tiền tảo lục - Prochlophyceae
Năm 1975 Ralph Lewin thông báo đã phát hiện một nhóm tảo tiền nhân
không có phycobilin và có sắc tố chlorophyll b. Tảo này giống với tảo lam ở các
đặc điểm cấu trúc tế bào và sinh hoá, chỉ khác là thiếu các phycobilisome trên 11
mặt thylakoid và tảo này được đặt tên là Prochloron. Tảo lớp này không có dị bào.
Chi Prochloron (hình 6) sống cộng sinh ở Hải tiêu; Prochloron chỉ gặp ở
các vùng nước ven biển với nhiệt độ biến động từ 210C- 31 C 0 và giảm quang
hợp ở nhiệt độ dưới 200C. Một số Prochloron có khả năng cố định nitơ nhưng
chỉ trong điều kiện sống cộng sinh với Hải tiêu.
Hình 2.3. Prochloron didemni
Prochloron, ngoài chlorophyll a còn có chlorophyll b và thiếu
phycobiliprotein. Chính vì thế mà tảo này có màu lục, màu này không có ở tế bào tảo lam.
Chi Prochlorococcus sống phiêu sinh, dạng tế bào hình cầu không có vỏ
bao; chi Prochlorothrix dạng sợi, có chứa túi khí và sống ở nước ngọt.
II. Ngành tảo đỏ - RHODOPHYTA
1. Tính chất đặc trưng của ngành
Tảo đỏ khác biệt với các ngành
tảo có nhân thật khác ở điểm tế bào
không mang roi. Hệ thống quang hợp
gồm chlorophyll a, β-carotene và nhóm phycobilin gồm phycoerythrine,
phycocyanin và allophycocyanin. Ba
sắc tố này tập trung trong thể
phycobilisome nằm ở bề mặt ngoài của
thyllakoid. Đôi khi sắc tố chlorophyll d
cũng gặp ở tảo đỏ nhưng chức năng
của nó trong quá trình quang hợp chưa
được khẳng định. Sản phẩm dự trữ là
một carbohydrate tương tự như
glycogen hoặc amylopectin - một dạng tinh bột.
Ở đa số tảo đỏ có thành tế bào cấu tạo bởi chất cellulose, nhưng cũng có
loài thành tế bào cấu tạo bởi mannan (một polymer của mannose) hoặc xylan
(polymer của xylose). Chất keo nhầy gặp ở thành tế bào của tảo đỏ gồm agar- 12
agar và caragheen. Đây là polymer của đường galactose và nó có giá trị thương
phẩm quan trọng nhờ ở tính ổn định và tính chất keo (gel).
Tế bào tảo đỏ đặc trưng có một đến nhiều nhân. Một không bào lớn nằm ở
vùng trung tâm với một hoặc vài lục lạp nằm ở lớp tế bào chất bao quanh.
Thylakoid nằm rời nhau, bề mặt mang các hạt phycobilisome - cấu trúc tiếp nhận
ánh sáng chính của tế bào. Năng lượng ánh sáng sẽ được chuyển đến chlorophyll
nằm ở bên trong thylakoid:
Phycoerythrin==>phycocyanin==>allophycocyanin==>chlorophyll a.
Khi phycoerythrin ưu thế, tảo có màu đỏ, tuy vậy màu sắc của tảo đỏ rất thay đổi.
Lục lạp tảo đỏ có cấu trúc vỏ gồm hai màng và được cho rằng bắt nguồn từ
thể tảo lam cộng sinh. Lục lạp chứa các hạt tạo bột, các hạt tinh bột nằm trong tế
bào chất. Đa số tảo đỏ có cấu trúc cơ thể đa bào và sinh trưởng bằng phân cắt dinh dưỡng.
Trong quá trình phân chia nguyên phân vỏ bao nhân vẫn tồn tại. Một số ít
tảo đỏ dạng đơn bào hoặc dạng phiến cấu trúc theo kiểu mô giả còn lại là tảo
dạng sợi. Một số tảo dạng sợi mang nhánh tự do, và đa phần tảo có cấu trúc tản
kiểu dáng mô giả pseudoparenchymatous trong đó khó phân biệt được từng sợi riêng lẽ.
2. Phân loại và phân bố
Tảo đỏ được phân làm hai lớp: Bangiophyceae và Floridiophyceae.
Tảo đỏ có đặc điểm nổi bật là thành tế bào hình thành các điểm liên kết
giữa các tế bào kế nhau. Dựa vào đặc điểm của điểm liên kết này để phân tảo đỏ làm hai dưới lớp.
Trong phân chia tế bào ở lớp Floridiophyceae sự hình thành vách ngăn
không hoàn toàn, giữa trung tâm của vách ngăn ngang hai tế bào kế cận còn lại
một điểm liên kết có một nút đậy. Điểm liên kết thứ cấp có thể được hình thành
ở các tế bào không có quan hệ. Tất cả các loài thuộc Floridiophyceae đều có cấu
trúc đặc trưng về nút đậy. Tuy nhiên, vẫn phát hiện nút đậy ở một số giai đoạn
của lớp Bangiophyceae. Nút đậy được hình thành trong khi phân chia tế bào chất. 13
Tảo Floridiophyceae thường sinh trưởng ở đỉnh, tế bào có nhiều sắc thể
nhỏ và chu trình sống phức tạp. * Lớp Bangiophyceae
Những đặc điểm chính của lớp
Tản tương đối đơn giản: đơn bào, dạng sợi hoặc phiến, tế bào đơn nhân.
Sinh trưởng bằng sự phân cắt của tế bào đỉnh hoặc của tế bào xen.
Có hoặc không có cấu trúc điểm ở thành tế bào.
Sắc thể thường hình sao và nằm ở trung tâm tế bào. Tuy nhiên, một vài
loài có sắc thể nằm ở bên hoặc sắc thể hình đĩa.
Các loài sinh sản hữu tính có chu trình sống đơn lưỡng bội đồng hoặc dị
hình luân phiên thế hệ. Ở dạng sinh sản hữu tính, thư quả gần như không khác
mấy so với tế bào dinh dưỡng về hình dạng, thiếu thư mao. Thư quả không tạo ra
thể quả bào tử lưỡng bội.Không có tứ bào tử.
Chu trình sống của Porphyra
Phần đứng của tản dạng lá thường cấu tạo bởi 1 lớp tế bào. Phiến thường
gấp nếp không đều hoặc xẻ mép phiến. Tản bám vào giá thể bằng rể giả. Mỗi tế
bào chứa 1 sắc thể hình sao nằm ở trung tâm tế bào và chứa hạt tạo bột. 14
Phần lớn loài là lưỡng tính, nhưng cũng có một số chỉ gặp toàn cây đực.
Các tế bào dinh dưỡng biến đổi thành túi tinh tử (túi giao tử đực) chứa nhiều tinh tử.
Thư quả hầu như không phân biệt với các tế bào dinh dưỡng xung quanh.
Ngoại trừ chúng có mang một mấu lồi dạng núm.
Tinh tử được dòng nước đưa đến đậu vào bề mặt tản. Nhân tinh tử được
chuyển qua một khe nhỏ vào bên trong thư quả, tại đây nhân tinh tử và nhân tế
bào trứng phối hợp. Sau khi thụ tinh, thư quả phân chia cho ra 4, 8, 16, 32 tế bào
lưỡng bội - bào tử quả (carpospore).
Mỗi một bào tử quả phát triển thành hệ thống sợi nẩy nhánh mà chúng có
thể mọc luồn vào trong cấu trúc vỏ của nhóm hai mảnh vỏ. Tảo dạng sợi nẩy
nhánh này đầu tiên được phát hiện như là 1 loài tảo đỏ mới và được đặt tên là Conchocelis rosea.
Chính vì vậy, pha dạng sợi của Porphyra còn gọi là pha conchocelis. Cơ
thể hình thành quả bào tử và pha conchocelis là lưỡng bội. Bào tử conchocelis
hình thành từ conchocelis sẽ phát triển thành cá thể Porphyra hình phiến. Cây
Porphyra non có thể sinh sản bằng đơn bào tử ở mép trên của phiến.
Người ta cho rằng pha Porphyra tồn tại theo mùa đông hằng năm, trong
khi pha conchocelis sống lâu năm, nhờ khả năng sống được trong điều kiện mùa hè.
Theo Magne, bào tử conchocelis và pha conchocelis là lưỡng bội, cây
Porphyra đơn bội. Quá trình giảm phân xảy ra khi conchospore nẩy mầm. Chu
trình sống của Porphyra hai kỳ đơn lưỡng tính. * Lớp Floridiophyceae
Những đặc điểm chính của lớp: Tản thường có dạng sợi nẩy nhánh. Tuy
nhiên, thường kết nhau thành cấu trúc mô giả. Tản hình ống dẹt hoặc hình lá. Tế
bào đơn nhân hoặc đa nhân. Sinh trưởng chủ yếu là do tế bào đỉnh, một số có sự
phân cắt của tế bào xen. Thành tế bào luôn có cấu trúc điểm thông liên kết. Sắc
thể ở phía bên, có dạng dải hoặc đĩa, không có hạt tạo bột. Tuy nhiên cũng gặp
một số loài sắc thể dạng sao và có hạt tạo bột ở trung tâm và nằm ở giữa tế bào.
Các loài sinh sản hữu tính có chu trình sống đơn lưỡng tướng đồng hoặc
dị hình, hiếm khi có chu trình đơn tướng. Thư quả khác biệt rõ với tế bào dinh
dưỡng vì nó phát triển 1 thư mao có chức năng "hứng" lấy tinh tử và tiếp nhận
nhân của tinh tử. Sợi sinh quả bào tử đã thụ tinh sẽ phát triển thành thể quả bào
tử lưỡng bội. Thể này sẽ tạo ra bào tử quả lưỡng bội và bào tử này phát triển
thành thể tứ bào tử lưỡng bội. Thể tứ bào tử lưỡng bội hình thành bào tứ bào tử
đơn bội. Tứ bào tử phát triển thành cây giao tử đơn bội.
Tảo lớp này có chu trình sống phức tạp. Ngoài chu trình hai pha đơn bội
và lưỡng bội xen kẽ nhau, tảo còn có chu trình ba pha gồm thể giao tử, thể quả
bào tử và thể tứ bào tử. Thể giao tử đơn bội sản sinh giao tử không mang roi.
Sinh sản hữu tính noãn giao. Thể quả bào tử lưỡng bội phát triển từ hợp tử trên
cây giao tử cái và tạo ra bào tử để hình thành nên thể tứ bào tử. Phân chia giảm 15
nhiễm xảy ra khi hình thành tứ bào tử và tứ bào tử phát triển thành thể giao tử.
Vì thế chu trình sống của tảo có hai pha lưỡng bội: thể quả bào tử và thể tứ bào
tử. Thể tứ bào tử và thể giao tử là những pha sống độc lập (sống tự do), trong khi
thể quả bào tử gắn liền với thể giao tử.
Trong chu trình sống của tảo Floridiophyceae, thể giao tử đực và cái sống
tự do. Thể giao tử đực hình thành nên tinh tử trong nhánh sinh tinh. Tinh tử
không có sắc tố. Thể giao tử cái hình thành nên một nhánh đặc biệt gọi là nhánh
mang thư quả (carpognium) trong đó tế bào tận cùng giữ chức năng như một túi
trứng. Thư quả có cấu trúc dạng một chiếc độc bình với phần đỉnh kéo dài được
gọi là thư mao (trichogyne) thường nhô ra khỏi bề mặt của thể giao tử cái; phần
còn lại của thư quả nằm trong tản. Nội chất bên trong thư quả được xem như là
tế bào trứng. Khi tinh tử được nước đưa đến đậu vào thư mao, nhân tinh tử sẽ
xâm nhập qua thành thư mao và đi xuống phối hợp với tế bào trứng. Thể quả bào
tử có thể phát triển trực tiếp từ thư quả đã thụ tinh hoặc nhân hợp tử di chuyển
qua một trợ bào (auxiliary cells) để hình thành nên thể quả bào tử. Thể quả bào
tử gồm các sợi nẩy nhánh được gọi là sợi sản bào (gonimoblast) gồm các tế bào
lưỡng bôị được sản sinh từ tế bào hợp tử. Thể quả bào tử có thể là một cụm sợi
kết dính được coi như là một cấu trúc đặc thù gọi là quả túi (cystocarp) nằm
trong thể giao tử cái, hoặc là cụm sợi phát triển cùng với các sợi của thể giao tử.
Một số tế bào của thể giao tử có thể tạo nên một lớp vỏ quả (pericarp) bao quanh
thể quả bào tử để cung cấp chất dinh dưỡng cho thể quả bào tử. Tại đỉnh của các
sợi sản bào, quả bào tử được hình thành.
Quả bào tử nẩy mầm cho ra thể tứ bào tử (tetrasporphytes) lưỡng bội. Khi
thành thục, thể này hình thành nên các túi tứ bào tử đơn bào cho ra tứ bào tử đơn
bội. Các bào tử khi được phóng thích vào nước sẽ tách rời nhau, bám vào giá thể
và nẩy mầm cho ra thể giao tử mới.
Các kiểu chu trình sống gặp ở lớp tảo Floridiophycidae:
- Chu trình sống kiểu Polysiphonia
Trong chu trình sống của Polysiphonia cả thể giao tử lẫn thể tứ bào tử
tương tự về hình dáng. Nhánh mang tinh tử không màu và cho ra một cụm tinh
tử; thể giao tử cái hình thành nhánh mang thư quả gồm 4 tế bào trong đó tế bào
tận cùng là thư quả. Nhân của hợp tử sẽ chuyển vào một trợ bào nằm cạnh thư
quả và thể quả bào tử được bao quanh bởi một lớp vỏ quả rất phát triển
(pericarp). Thể tứ bào tử đồng dạng hình thái với thể giao tử và mang tứ bào tử ở đỉnh nhánh.
- Chu trình sống của Audouinella investeins
Tảo Audouinella investeins sống bì sinh trên tảo đỏ kích thước lớn
Batrachospermum. Người ta cho rằng kiểu vòng đời của loài này là kiểu cơ bản
mà từ đó các loài khác sẽ phát sinh. Cơ thể giống khối cầu lông nhỏ (dạng trái
banh nhỏ) gồm một cụm lông tơ được cấu tạo bởi các sợi tế bào nẩy nhánh. Tản
có thể là cây giao tử hoặc cây bào tử. Cây giao tử, mang cả túi tinh lẫn túi trứng.
Thư quả gồm 2 phần: phần gốc phình ra chứa nhân giao tử cái, phần trên là mấu
lồi dài, mảnh, không màu gọi là thư mao (trichogyne). Thư quả (carpogonium) 16
có cuống hoặc không cuống. Tinh tử được nước đưa đến đậu vào thư mao, nhân
tinh tử xâm nhập vào thư mao và chuyển xuống thư quả để phối hợp với nhân
trứng. Sau khi thụ tinh, thư quả phân chia tạo nên một hệ thống các sợi sản bào
(gonimoblast). Toàn bộ hệ thống sợi sản bào được gọi là thể quả bào tử
(carposporophyte). Bào tử quả nảy mầm phát triển thành cây tứ bào tử
(tetraspophyte) có hình dạng tương tự với cây giao tử đơn bội. Cây tứ bào tử khi
trưởng thành sẽ phân chia giảm nhiễm cho ra tứ bào tử đơn bội trong các túi tứ
bào tử. Tứ bào tử phát triển cho ra cây giao tử một vòng đời được khép kín.
- Chu trình sống kiểu Bonnemaisonia.
Chu trình Bonnemaisonia, thể tứ bào tử và thể giao tử khác nhau về hình
thái. Thường thể giao tử có tản cấu trúc dạng mô giả trong khi thể tứ bào tử có
dạng sợi đơn giản hay dạng mào gà. Do hình dáng khác biệt giữa hai thể này nên
chúng được xem là hai chi khác nhau. Nhưng khi nuôi cấy thì rõ là một loài gồm hai pha.
- Chu trình sống kiểu Palmaria.
Tản gồm các nhánh dạng phiến, màu đỏ đậm. Trong quần thể tự
nhiên chỉ có thể giao tử đực và thể tứ bào tử. Van der Meer và Todd (1980)
giải thích rằng tứ bào tử hình thành nên thể giao tử đực và cái với số lượng
bằng nhau, nhưng thể giao tử nhỏ, không lớn hơn 1mm và thành thục trong
vòng vài ngày trong khi thể giao tử đực lớn hơn phát triển từ 9 đến 10 tháng
để hình thành tinh tử. Vì vậy, tinh tử của thế hệ giao tử đầu tiên, thể giao tử
được hình thành từ đợt trước, tiến hành thụ tinh với giao tử của thế hệ giao
tử cái mới được hình thành. Tứ bào tử thể phát triển trực tiếp từ thư quả
và sống ngay trên thể giao tử cái. Vì thế vắng đi giai đoạn hình thành quả bào tử.
III. Ngành tảo dị roi - HETEROKONTOPHYTA
Ngành gồm các đại diện có tế bào mang hai roi không giống nhau, một roi
dài và một roi ngắn. Roi dài có phủ hai hàng lông tơ hướng về phía trước và roi 17
ngắn trơn hướng về sau. Lông phủ trên roi dài gọi là lông ống (mastigoneme) có
cấu tạo bởi ba phần: phần gốc, phần cán hình ống và một hoặc nhiều lông tơ nhỏ ở đỉnh cán.
Ngành này được cấu tạo bởi một nhóm tự nhiên và phân làm nhiều lớp
mặc dầu trong đó gồm cả tảo có cấu trúc đơn bào nhỏ như tảo silic đến tảo nâu
có kích thước tản đa bào lớn hằng chục mét. Tuy vậy giữa các nhóm tảo của
ngành vẫn thể hiện tính cận thân rất cao. Ngành gồm ít nhất 9 lớp: 1. Lớp Chrysophyceae 2. Lớp Parmophyceae 3. Lớp Sarcinophyceae 4. Lớp Xanthophyceae 5. Lớp Eustigmatophyceae 6. Lớp Bacillariophyceae 7. Lớp Rhaphidophyceae 8. Lớp Dictyophyceae 9. Lớp Phaeophyceae
Lớp 1, 4, 5 chủ yếu ở nước ngọt; lớp 2, 3, 8, 9 chủ yếu ở biển; lớp 6, 7 ở
nước ngọt và biển. Lớp 1 đến lớp 8 có cơ thể đơn bào hoặc đa bào đơn giản, lớp
9 tản cấu trúc đa bào với kích thước lớn.
Hiện nay người ta thừa nhận ngành Heterokontophyta bao gồm không chỉ
là những tảo quang hợp mà cả một số sinh vật không màu (sống dị dưỡng) động
vật nguyên sinh và cả nấm.
5.1. Lớp tảo vàng ánh - Chrysophyceae
Gồm những tảo đơn bào và tập đoàn có hoặc không có roi. Tuy nhiên
cũng có một số ít cấu trúc đa bào. Roi gắn ở đỉnh tế bào. Bộ phận cảm nhận ánh
sáng gồm một phần phồng lên của gốc roi ngắn và điểm mắt nằm trong sắc thể.
Bào tử nghỉ được hình thành trong tế bào. Các loài sinh sản hữu tính có lẽ
có chu trình sống đơn tướng, hợp tử trải qua giai đoạn sống nghỉ dưới dạng một
túi nghỉ (hypozygote) và có thể giảm phân khi nảy mầm.
Các đại diện của lớp này chủ yếu sống phiêu sinh ở nước ngọt, dạng đơn bào hay tập đoàn.
Tảo có màu vàng ánh (màu vàng kim) do các sắc tố phụ zeaxanthin,
antheraxanthin, volaxanthin, diatoxanthin, didinoxanthin lấn át sắc tố
chlorophyll a. Sản phẩm dự trữ (sản phẩm quang hợp) là chrysolaminarin
(leucosin), cũng có thể gặp các hạt dầu.
Lớp tảo này có khoảng 200 chi và 1000 loài. Tảo sống phiêu sinh ở môi
trường nước ngọt, một số ít sống ở môi trường nước lợ và nước mặn. Tảo vàng
ánh phát triển ưu thế ở các hồ nghèo dinh dưỡng có pH từ acid đến trung tính và
thích sống ở các vực nước lạnh. Ở các hồ giàu dinh dưỡng có pH trên 7,5 thường
nghèo nhóm tảo này. Tảo vàng ánh giữ vai trò quan trọng trong quyết định năng
suất sơ cấp của đại dương. 18
Chi Ochromonas là đại diện tiêu biểu của lớp, tảo dạng đơn bào trần và có
khả năng chuyển động kiểu amip. Chi này có khoảng 80 loài. Tế bào
Ochromonas mang hai roi không bằng nhau về kích thước và cấu trúc. Trong đó
roi dài hướng về phía trước có phủ hai hàng lông; roi ngắn láng, hướng về phía sau tế bào.
Ochromonas hình thành bào tử nghỉ bên trong tế bào dinh dưỡng. Đây gọi
là quá trình nội sinh bào tử nghỉ đặc trưng của lớp. Ở Ochromonas, sinh sản hữu
tính chưa được phát hiện. Tuy nhiên, một số đại diện của lớp như Dinobryon có
quá trình sinh sản hữu tính đẳng giao.
Cơ thể của tảo Chrysophyceae thuộc các kiểu sau:
- Dạng đơn bào, mang roi: Ochromonas (hình 16a), Dinobryon (hình 16b), P seudokephyton
- Dạng tập đoàn mang roi: Synura (hình 16c) - Dạng hạt: Chrysophaeta - Dạng amip: Rhizochrysis - Dạng palmella - Dạng sợi - Dạng tản
5.2. Lớp tảo vàng - Xanthophyceae
Lớp gồm các đại diện ở nước ngọt, chỉ một ít ở biển. Lớp đặc trưng bởi tế
bào chuyển động có roi dài gắn phía trước có phủ lông tơ, roi gắn phía sau trơn
giữ chức năng quạt nước.
Lục lạp chứa chlorophyll a và c, nhóm sắc tố phụ gồm carotene,
vaucheriaxanthin, diatoxanthin, diadinoxanthin và heteroxanthin; thiếu sắc tố
fucoxanthin nên tế bào có màu vàng nhạt.
Điểm mắt luôn ở trong sắc thể và được bao quanh bởi hai lớp màng của
mạng lưới nội sinh chất lục lạp, màng bên ngoài của lưới nội sinh chất lục lạp
kéo dài tiếp nối với màng nhân.
Lớp tảo vàng có khoảng 100 chi với 600 loài, những loài tảo đơn bào sống
phiêu sinh ở các hồ nước ngọt, một số ít loài ở biển và một số sống ở đất, đất đầm lầy.
Rất khó phát hiện tảo Xanthophyceae bởi vì chúng ít khi phát triển phong
phú ở bất cứ khu cư trú nào, ngoài trừ một số loài dạng sợi thuộc chi Tribonema
và các loài dạng ống Vaucheria. 19
5.3. Lớp tảo Silic - BACILLARIOPHYCEAE
Tảo silic là những tảo đơn bào, tập đoàn, sống phù du và sống bám; tảo
silic có thể sống quang dưỡng, tự dưỡng và dị dưỡng. Thành tế bào cấu tạo như
một chiếc hộp gồm hai nắp lồng vào nhau. Nắp trên (epitheca) gồm mặt vỏ trên
và dải bên trên; nắp dưới gồm dải bên dưới và mặt vỏ dưới; dải bên dưới và dải
bên trên kết hợp lại thành đai tế bào (girdle). Vì thế tế bào có thể nhìn thấy ở
phía mặt đai hay mặt vỏ. Vỏ tế bào được cấu tạo bằng chất silic. Tế bào tảo silic
rất đa dạng, chúng được xem là vật trang sức của thế giới sinh thuỷ sinh vật biển.
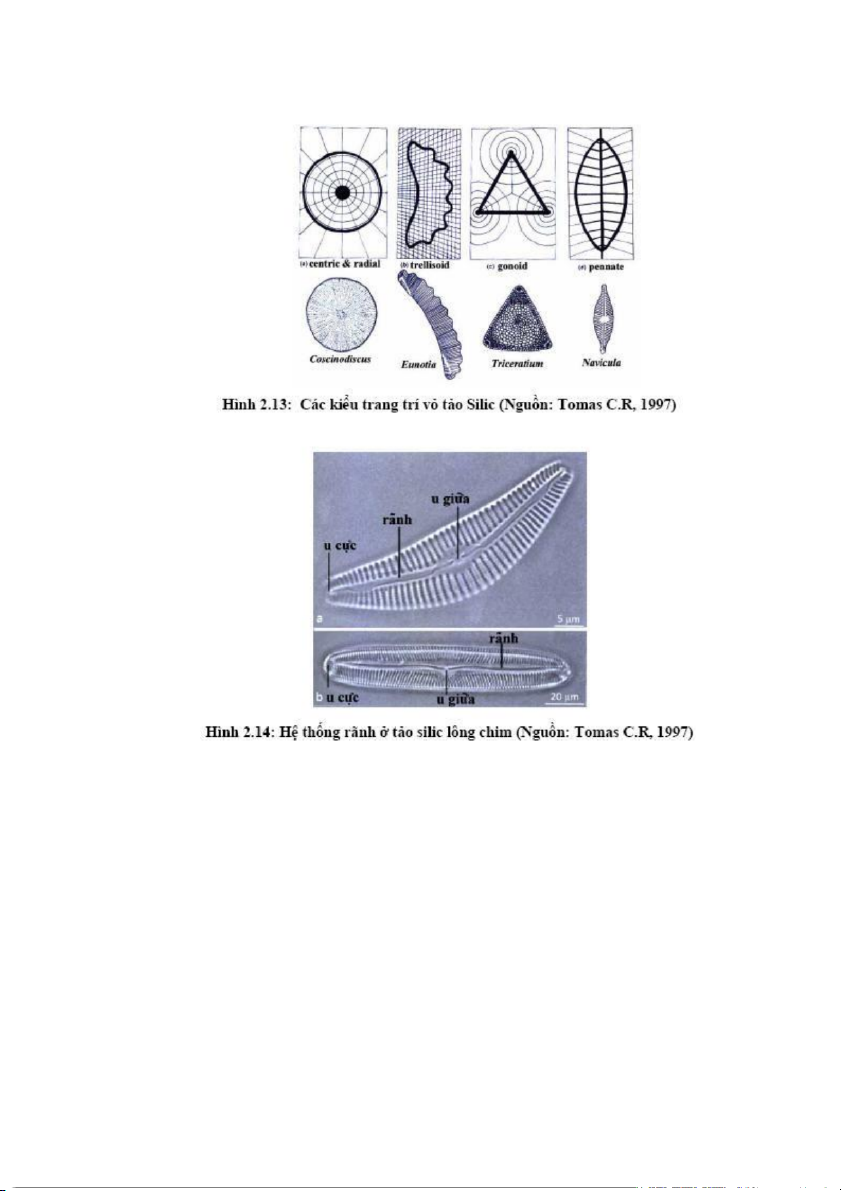
Mặt vỏ tảo mang những cấu trúc nhỏ sắp xếp theo các kiểu nhất định. Có
thể chia làm các kiểu cơ bản: i) kiểu centric, các cấu trúc sắp xếp dựa vào điểm
trung tâm của vỏ tế bào, cấu trúc vỏ đối xứng qua tâm điểm này; ii) kiểu
trelliosid, các cấu trúc sắp xếp đồng nhất trên bề mặt và không dựa vào tâm
điểm hay một đường thẳng; iii) kiểu gonoid, cấu trúc sắp xếp làm nổi bậc lên
các góc; iv) kiểu pennate, các cấu trúc xếp đối xứng hai bên, đối xứng qua
đường thẳng ở giữa tế bào. (hình 2.13)
Một số vỏ tảo silic lông chim có hệ thống rãnh gồm đường nứt dọc ở giữa
vỏ bị gián đoạn bởi một u lồi ở trung tâm vỏ (central nodule) và hai nốt phồng ở 20
2 phần cuối của rãnh được gọi là các u lồi ở cực tế bào (polar nodule). (hình 2.14)
Một số tảo không có rãnh nhưng ngay phần trung tâm có một phần không
cấu trúc chạy dọc tế bào được gọi là rãnh giả (pseudoraphe) (cấu trúc thoạt nhìn
giống như một rãnh). Rãnh không phải là một đường nứt đơn giản ở thành tế bào
mà là một khe hình chữ S trong đó phía ngoài rộng và ở phần giữa hẹp lại. 21
Phân chia tế bào ở tảo silic:
Sinh sản dinh dưỡng bằng phân đôi tế bào. Trước khi phân chia, nội
dung tế bào trương phồng lên đẩy tách hai nắp vỏ. Nhân phân chia và tiếp
theo là phân chia nguyên sinh chất bằng sự thắt vào của màng tế bào. Thể
golgi tạo ra các túi nằm ngay dưới màng tế bào (Stoermer et all, 1965) sẽ
hoà nhập lại với nhau tạo nên túi ngưng kết silic (Li et al., 1989). Các túi
này sẽ mở rộng dần và định hình hình dạng cho một vỏ mới. Hai túi được
tạo ra ở mỗi tế bào và sự ngưng kết chất silic trong mỗi túi sẽ hình thành
nên nắp vỏ dưới mới.
Mỗi tế bào con sẽ nhận một nắp của tế bào mẹ làm nắp trên còn nửa nắp
mới được hình thành luôn luôn là nắp dưới. Nắp tế bào mẹ chỉ có khả năng sử
dụng cho tế bào con trong một số lần. Chẳng hạn, ở Stephanopyxis (hình 2.15) ,
nắp tế bào mẹ chỉ dược dùng lại cho 6-8 thế hệ.
Kết quả của một lần phân chia tế bào sẽ tạo nên một tế bào con có kích
thước bằng tế bào mẹ và một tế bào có kích thước nhỏ hơn.
Do sẽ có một nửa số tế bào con giảm kích thuớc sau mỗi lần phân chia nên
trong quần thể tảo silic sẽ có một số tảo giảm dần kích thước trong tiến trình
phân chia dinh dưỡng tế bào. Khi tế bào giảm đến một kích thước tối thiểu nhất
định, khoảng 1/3 kích thước tối đa của tảo thì tế bào tiến hành quá trình khôi
phục kích thước bằng hình thành bào tử sinh trưởng thông qua sinh sản hữu tính.
Những tế bào kích thước quá nhỏ (nhỏ hơn 1/3 kích thước tối đa của tảo) không
hình thành được bào tử sinh trưởng thì sẽ tiếp tục phân chia cho đến khi chết.
Hình thành bào tử sinh trưởng ở tảo silic
Hình thành bào tử sinh trưởng luôn gắn liền với sinh sản hữu tính. Vì thế,
sinh sản hữu tính của tảo silic sẽ xảy ra khi: i) tế bào phải đạt một kích thước tối
thiểu nhất định (thường bằng khoảng 1/3 kích thước cực đại của tế bào dinh
dưỡng); ii) một số yếu tố môi trường thoả mãn cho tảo phát triển: nhiệt độ, ánh
sáng, muối dinh dưỡng, chất hữu cơ... Sinh sản hữu tính để hình thành nên bào
tử sinh trưởng được coi là một cơ chế thứ hai để khôi phục lại kích thước nguyên
bản. Bào tử sinh trưởng được hình thành bằng sự phối hợp của hai giao tử. Ở tảo
silic trung tâm tế bào giao tử đực chuyển động bằng roi, tế bào giao tử cái là tế
bào trứng; ở tảo silic lông chim, tế bào giao tử không mang roi. Cũng có trường
hợp, bào tử sinh trưởng được hình thành không qua sự phối hợp giao tử (tuy
nhiên nhiều trường hợp cần phải được xem lại). Có thể khái quát quá trình hình
thành bào tử sinh trưởng như sau: Nguyên sinh chất thoát khỏi vỏ tế bào mẹ, lớn 22
dần lên và tạo ra một màng bằng chất silic đặc biệt gọi là perizonium gồm nhiều
dải xếp chồng lên nhau. Tế bào dinh dưỡng với kích thước hình thái bình thường
sẽ được hình thành trong màng perizonium này và tế bào sẽ phá vỡ màng thoát
ra ngoài. Trầm tích chứa nhiều màng perizonium và vỏ tảo silic thường thấy ở
các vùng vào thời điểm cuối của hiện tượng tảo silic nở hoa.
Ở tảo silic lông chim, sinh sản hữu tính theo kiểu tiếp hợp của hai giao tử.
Ở Pinnularia, Navicula hai tế bào tiến đến bên nhau và cùng được bao trong một
bao nhầy. Nhân của mỗi tế bào thực hiện hai lần phân chia cho ra 4 nhân con
trong đó có một nhân bị huỷ sau mỗi lần phân chia. Kết quả mỗi tế bào có hai
nhân, mỗi nhân cùng với một phần nguyên sinh chất giữ chức năng là giao tử.
Một giao tử của tế bào thứ nhất sẽ di chuyển sang phối hợp với giao tử bị động
của tế bào thứ hai và giao tử còn lại sẽ bị động chờ giao tử của tế bào thứ hai
sang phối hợp. Kết quả của quá trình này cho ra một bào tử sinh trưởng ở một tế
bào. Bào tử sinh trưởng sẽ lớn dần lên để cho ra một tế bào mới.
Vậy chu trình sống của tảo silic là một kỳ lưỡng bội.
Bào tử và tế bào nghỉ
Bào tử và tế bào nghỉ cho phép tảo silic tồn tại trong một thời kỳ mà điều
kiện môi trường bất lợi như có tuyết phủ, suy kiệt chất dinh dưỡng hay sự phân
tầng cột nước và sau đó khi môi trường đã được cải thiện tảo sẽ phát triển trở lại.
Bào tử và tế bào nghỉ góp phần quan trọng trong chuyển tải carbon hữu cơ
và silicate xuống trầm tích đáy. Bào tử của tảo silic Chaetoceros có thể chiếm
hơn 50% của trầm tích đáy của các vùng ven biển phía bắc Thái Bình Dương.
3. Phân loại: Lớp tảo silic được chia thành 2 bộ là bộ tảo silic trung tâm
(Centrales) và bộ tảo silic lông chim (Pennales).
-Bộ Centrales: tế bào đơn độc hoặc kết thành chuỗi, dạng hình hộp tròn,
đĩa, bầu dục, nhiều cạnh hoặc dạng ống... Vân hoa trên mặt vỏ sắp xếp theo dạng
đối xứng toả tia. Không có cấu tạo rãnh dài hoặc rãnh giả, không có khả năng di
động. Chi điển hình: Coscinodiscus, Thallasiosira, Chaetoceros, Stephanopyxis... 23
- Bộ Pennales: Tế bào hình que thẳng, cong hình chữ S, hình lưỡi liềm,
hình nêm, bầu dục, cong yên ngựa ... Tế bào có rãnh, có hoặc không có khả năng
di chuyển. Vân hoa trên mặt vỏ sắp xếp theo dạng đối xứng hai bên đối với trục
dài như dạng lông chim. Chi điển hình: Navicula, Pleurosigma, Diploneis, Nitzschia, Pinnularia ...
Chi Navicula: tế bào hình mũi mác, ellip ... hai đầu nhọn hoặc tròn, vùng
trục rõ ràng, rãnh và nốt cực phân biệt. Vân thẳng, cong hoặc toả tia.
Chi Diploneis: tế bào hình bầu dục có hoặc không có eo thắt ở giữa, đỉnh
tròn hoặc nhọn. Nốt giữa to, hình vuông, có các gốc kéo dài thành nhánh song
song với rãnh dài. Các lỗ vân sắp xếp thành các đường vân cong theo chiều cong
của mép vỏ và thẳng dần ở phần giữa tế bào. Hai bên rãnh có hai tuyến trơn dài.
5.4. Lớp tảo nâu - PHAEOPHYCEAE
Tảo nâu có khoảng 265 chi và 1500-2000 loài. Tảo nâu chủ yếu sống ở
biển. Từ những năm 1920, tảo nâu đã được sử dụng làm nguyên liệu tươi để
sản xuất alginat. Những tảo nâu được tập trung khai thác là Macrocytis,
Nereocystis, Laminaria, và Ascophyllum.
Lớp tảo nâu khác biệt rõ với các lớp còn lại của ngành Heterokontophyta.
Tất cả các loài tảo nâu có cấu trúc đa bào. Hình dạng tản thay đổi từ dạng sợi có
nhánh kích thước hiển vi đến tản có kích thước lớn hằng mét. Chỉ có tế bào sinh
sản có roi và roi gắn phía bên tế bào. Bộ phận cảm nhận ánh sáng gồm một phần
phình lên của roi sau và một điểm mắt chứa trong lục lạp. Vùng chuyển tiếp của
roi thiếu bộ phận xoắn. Thành tế bào cấu tạo bởi một hệ thống vi sợi cellulose
được làm vững chắc bởi calcium alginat cùng với phần cơ chất nhầy không định
hình. Lục lạp có lưới nội sinh chất lục lạp bao quanh và có thể kéo dài tiếp xúc 24
với màng nhân. Thylakoid ngoại vi nằm bên dưới vỏ lục lạp. Lục lạp chứa
chlorophyll a, c1, c2. Sắc tố fucoxanthin lấn át chlorophyll làm cho tảo có màu
nâu. Sản phẩm quang hợp là chrysolaminarin. Tảo nâu có chu trình sống hai kỳ
đơn lưỡng bội đồng hình hoặc dị hình và chu trình một kỳ lưỡng bội. Tế bào sinh
sản được hình thành trong cơ quan sinh sản một ngăn và nhiều ngăn. Các giao tử
được hình thành trong túi giao tử một ngăn và nhiều ngăn; bào tử được hình
thành trong túi bào tử một ngăn và nhiều ngăn trong đó túi bào tử một ngăn cho
ra bào tử đơn bội và túi nhiều ngăn cho bào tử lưỡng bội.
Phần lớn tảo nâu có sinh sản hữu tính đều có chu trình sống hai kỳ đơn
lưỡng tướng sinh và có thể đồng hình hay dị hình luân phiên thế hệ. Ỏ chu trình
luân phiên thế hệ dị hình, có thể thể giao tử đơn bội hay thể bào tử lưỡng bội tiêu
giảm. Thể giao tử hình thành nên túi giao tử nhiều ngăn (plurilocular
gametangia), mỗi ngăn sẽ cho ra một giao tử. Các giao tử có thể giống nhau hay
khác nhau về kích thước.
Nhiều tảo nâu tế bào giao tử cái không chuyển động. Thể bào tử cũng
mang túi bào tử một ngăn và nhiều ngăn. Ở túi bào tử một ngăn có sự phân chia
giảm nhiễm để hình thành nên bào tử đơn bội. Bộ Fucales đặc trưng với chu
trình sống 1 kỳ lưỡng bội trong đó sự phân bào giảm nhiễm xảy ra khi hình
thành giao tử. Ở đây cả giao tử đực và tế bào trứng được hình thành không chỉ
trong túi giao tử một ngăn mà cả túi giao tử nhiều ngăn.
Ectocarpus dạng sợi đơn giản không nẩy nhánh. Chu trình sống có luân
phiên thế hệ đồng hình (hơi dị hình). Thể giao tử đơn bội cho ra giao tử đơn bội
trong túi giao tử nhiều ngăn. Thể bào tử lưỡng bội cho ra bào tử đơn bội trong
túi bào tử một ngăn. Thường là dị giao phối, nhưng một số đại diện thực hiện noãn giao. 25
Tảo bộ Laminariales có chu trình sống dị hình luân phiên thế hệ rõ rệt với
thể bào tử có kích thước rất lớn, thể giao tử có kích thước nhỏ. Sự phân chia
giảm nhiễm xảy ra trong túi bào tử một ngăn. Thể giao tử cái sinh trứng và thể
giao tử đực sinh tinh trùng.
Bộ Fucales gồm tất cả các đại diện lưỡng bội và hầu hết có sinh sản hữu
tính noãn giao. Túi tinh và túi trứng phát triển trong các huyệt (conceptacle); các
huyệt tập trung lại trên một đế (receptacle) thường ở đỉnh của một số nhánh. Thể
lưỡng bội ở bộ này là thể bào tử. Bào tử thay vì được phóng thích nó lại nằm lại
trong túi bào tử có thực hiện phân chia giảm nhiễm và phát triển tại chỗ thành
thể giao tử đực và thể giao cái tiêu giảm. Các tế bào tiêu giảm này được xem như
sự tồn tại duy nhất của tản dinh dưỡng của thể giao tử. Với cách lý giải này, chu
trình sống của tảo bộ Fucales tương đồng với chu trình sống của thực vật có hoa.
Hình 2.23. Sơ đồ vòng đời của Fucus
IV. Ngành tảo roi bám - HAPTOPHYTA
Tảo đơn bào, cơ thể có dạng amip, dạng hạt, dạng pamella hoặc dạng sợi. 26
Tế bào mang hai roi bằng nhau hoặc không bằng nhau và roi không phủ
lông lơ. Tế bào có mang thêm một roi phụ gọi là roi bám
Sắc thể nằm bên trong của một nếp gấp của mạng lưới nội sinh chất. Sắc
thể chứa chlorophyll a, c (c1, c2, c3) không có chlorophyll b. Tảo có màu vàng
hoặc vàng ánh đó là do chlorophyll bị át bởi sắc tố phụ fucoxanthin. Một số sắc
tố carotenoid gồm carotein, diadinoxanthin và diatoxanthin. Sản phẩm dự trữ
quan trọng là chrysolaminarin. Ngoài ra paramylon cũng gặp ở một số loài. Bề
mặt tế bào đặc trưng nhờ có phủ những tấm vảy hoặc hạt có cấu tạo bằng
cellulose, ngoài ra còn có các tấm calci.
Chu trình sống của Haptophyta 2 kỳ dị hình luân phiên thế hệ, trong đó
giai đoạn mang hai roi 2n, giai đoạn sống phù du, xen kẽ với giai đoạn sợi n, giai
đoạn sống bám đáy. Haptophyta chủ yếu sống ở biển, chỉ số ít sống ở nước ngọt.
Hơn 25 năm qua, tảo này đã được nghiên cứu nhiều nhờ vào kính hiển vi
điện tử. Sự phát hiện lý thú nhất đó là các tấm vảy polysacharide.
Ngành này có khoảng 75 chi và 500 loài. Nó đóng vai trò quan trọng ở đại
dương tạo nên sức sản xuất sơ cấp của thuỷ vực.
Đa số tảo roi bám thuộc nhóm nanoplankton bao gồm cả nhóm plankton
(2-20μm chiều dài) và picoplankton (0,2-2μm chiều dài). Nhóm Haptophyta có
vảy calci hóa thạch được coi là nguồn cung cấp calci cho thuỷ vực. Đại diện:
Chrysochromulina, Prymnesium, Pleurochrysis.
V. Ngành Huyệt bào tảo - CRYPTOPHYTA
Ngành chỉ có một lớp Cryptophyceae. Phần lớn đại diện của lớp có dạng
đơn bào mang roi, mặc dù một số tảo ở một giai đoạn có cấu trúc tản dạng
pamella hoặc dạng bất động dính vào giá thể (sessile). Chỉ có một chi
Bjornbergiella có cấu trúc dạng sợi đơn giản.
Tế bào mang hai roi không bằng nhau, roi dài mang hai hàng lông ống
(mastigonemes), roi ngắn mang một hàng lông tơ. Tế bào có cấu trúc lưng bụng:
mặt lưng lồi, mặt bụng phẳng mang một rãnh dọc nông. (hình 2.25). Mặt đỉnh tế
bào cụt. Đỉnh trước của rãnh có một hầu sâu với thành hầu mang nhiều
trychocyts. Hai roi mọc ra phía trên và quanh bên phải của hầu. 27
Lục lạp được bao quanh bởi một nếp gấp của lưới nội sinh chất, lưới nội
sinh chất lục lạp nối tiếp với màng nhân. Khoảng giữa màng lưới nội sinh chất
lục lạp và lục lạp có lưới periplastidan. Khác với ngành Haptophyta và
Heterokontophyta, thylakoid ở lục lạp thường kết thành cặp tạo nên các tấm, bên
cạnh đó cũng gặp các thylakoid kết thành tấm lamell. Thylakoid ở tảo này cũng
dày hơn so với thylakoid của các ngành tảo khác. Các hạt tạo bột nhô ra ở mặt
trong của lục lạp. Nó có thể chứa các tấm lamelle, mỗi tấm có 1-2 thylakoid. Một
đặc điểm sự độc đáo của ngành tảo này là chất dự trữ tinh bột không được tích
luỹ trong lục lạp cũng như tế bào chất mà ở trong vùng trống periplastidan giữa
lưới nội sinh chất lục lạp và vỏ lạp.
Tế bào thường không có điểm mắt. Ở những đại diện có điểm mắt thì
điểm mắt ở dạng các giọt dầu hình cầu giống với Heterokontophyta. Điểm mắt
nằm ở giữa tế bào ngay trong lục lạp và không gắn liền với roi, đặc điểm này
khác với kiểu điểm mắt của Heterokontophyta. Tế bào được bao bởi màng
protein chắc thường tạo nên các tấm dạng hình chữ nhật. Mặt ngoài của màng
plasmalema được phủ bởi các tấm có cấu trúc tinh thể. Thể nhân nằm giữa lục
lạp và lưới nội sinh chất, có màng bao kép, có thủng lỗ và chứa DNA. 28
Sinh sản hữu tính đẳng giao, sự phối hợp nhân mới được quan sát gần đây
ở một loài. Ở một số loài chu trình sống gồm giai đoạn mang roi đơn bội và một
giai đoạn cơ thể lớn hơn vẫn đồng hình dáng và có roi nhưng lưỡng bội. Ở mỗi
giai đoạn, tế bào có thể cho ra cá thể khác. Chu trình sống hơi dị hình, đơn -
lưỡng bội. Đây là kiểu chu trình sống trong đó gồm giai đoạn mang roi đơn bội
xen kẽ với giai đoạn mang roi lưỡng bội.
Lớp tảo Cryptophyceae gặp ở cả môi trường nước ngọt và biển. Ngoài các
loài sống tự dưỡng, cũng gặp các loài sống dị dưỡng.
Lớp này gồm có 12 chi với khoảng 100 loài sống ở nước ngọt và 100 loài ở biển.
Các loài nước ngọt thường xuất hiện theo từng thời kỳ ở các ao nhỏ, các
vũng nước đọng đặc biệt ở những nơi giàu chất dinh dưỡng. Các loài sống ở biển
cũng xuất hiện phổ biến ở các ao đầm vùng triều hay các đầm nước lợ.
Cryptophyta chiếm giữ một phần quan trọng trong thành phần
nanoplankton (tảo phù du có đường kính 2-20μm). Vì thế đôi khi chúng trở nên
ưu thế trong nhóm gây nở hoa nước ở một giai đoạn nhất định vào mùa xuân ở
vùng trung tâm biển Bắc. Một số Cryptophyta có thể di chuyển theo cột nước
như Dinoflagellates. Trong điều kiện môi trường yên tĩnh chúng có thể sử dụng
roi để điều chỉnh vị trí phân bố theo chiều thẳng đứng. Các loài khác nhau chiếm
giữ một vị trí trí tối ưu khác nhau.
VI. Ngành tảo giáp- DINOPHYTA (ngành tảo 2 rãnh)
Tảo giáp gồm những cơ thể đơn bào, có nhân thật, chuyển động được nhờ
roi. Chúng thường sống đơn độc hoặc hình thành nên tập đoàn dạng chuỗi.
Thể màu hình bản, chứa chất diệp lục a và c, carotene ß và xanthophyll,
peridinin màu đỏ đậm, diadinoxanthyll, dinoxanthyll, neodinoxanthyll và
pyrrophyll màu nâu. Trên thể màu chứa một hoặc vài hạt tạo bột. 29
Sản phẩm quang hợp là các hạt dầu (lipit) thường nhuốm xanthophyll có
màu vàng hay đỏ và tinh bột được hình thành bên ngoài lục lạp đặc biệt bắt màu lam với iôt.
Cấu trúc vỏ tế bào:
Tảo giáp được chia thành hai nhóm chính
- Nhóm Dinokont: Cơ thể của nhóm này thường có dạng hai nửa hình cầu
úp lại, cơ thể có đai dọc (sulcus) và đai ngang (singulum). Gốc roi xuất phát từ
điểm giao nhau của đai ngang và đai dọc. Người ta chia nhóm này thành 2 nhóm:
+ Nhóm không có giáp: là những cơ thể với tế bào trần, cơ thể có
hai hình tròn với 2 lạp to mang những hạt tinh bột to và tròn.
+ Nhóm có giáp: Được bao phủ bên ngoài bởi các tấm cellulose.
- Nhóm Desmokont: Cơ thể được cấu tạo bởi hai nắp úp dẹt, không có đai
dọc và đai ngang. Các gốc roi ở phía đầu tế bào.
Màng tế bào hoặc bằng periplast (như ở Gymnodinium) hoặc có vỏ bằng
cellulose với các tấm giáp (plate). Trên bề mặt của các tấm giáp có thể nhẵn
hoặc được trang trí bằng các vân mịn hoặc thô và thường có nhiều lỗ nhỏ nhằm
làm tăng thêm diện tích bề mặt và dễ trôi nổi. Chất cấu thành áo giáp là một
polysaccharide, có thể là cellulose. Các tấm giáp này dính vào nhau, và làm
thành hai nửa, nửa trên gọi là nắp trên, nửa dưới gọi là nắp dưới. Nắp trên và nắp
dưới được nối với nhau bởi một vành đai ngang, còn đai dọc nằm ở nắp dưới của
tế bào, các vành đai này cũng được tạo bởi các tấm giáp, nhờ có các đai này mà
các tấm giáp mới có thể mở rộng ra và giúp cho tảo sinh trưởng. Số tấm và hình
dạng, kích thước của chúng được sử dụng như một đơn vị trong việc nghiên cứu về phân loại. 30 Cấu trúc của roi:
Roi của tảo giáp không đều nhau nằm trong rãnh ngang và rãnh dọc. Roi ở
rãnh ngang có tác dụng chuyển động vòng quanh còn roi ở rãnh dọc làm vận
động co rút. Các loài tiến hóa thấp, roi mọc ở đỉnh, còn những loài tiến bộ hơn
roi mọc ở sườn bụng. Cả hai roi đều có lông tơ mịn ở bên. Roi dọc mang một
hàng, roi ngang mang hai hàng lông tơ. Roi ngang chứa một thể xoắn, cả hai roi
đều xuất phát từ mặt bụng của tế bào.
Roi dọc mang các sợi mảnh (dài khoảng 0.5nm, dày 10nm), được xếp
thành hai hàng. Các sợi này có độ dày rất khác so với các sợi mastigonemes ở roi
của Heterokontophyta (dày khoảng 25nm). Roi dọc chạy dọc vùng dưới của
sulcus. Roi ngang chạy quanh tế bào ở đai ngang.
Các thể gốc của roi được giữ chặt trong tế bào bởi một hệ thống rễ. Sự
định hướng của các thể gốc, khác nhau rất nhiều giữa nhóm này với nhóm khác
trong ngành Dinophyta. Ở Gymnodinium, các thể gốc gần như có hướng đối
nhau (đối song song), trong khi ở Ceratium, chúng gần như nằm cùng phía bên
phải với nhau; một chi khác là Prorocentrum, các thể gốc gần như song song. Sinh sản:
Sinh sản sinh dưỡng bằng cách phân đôi tế bào.
Với các tế bào trần thì sự phân cắt của tế bào không phức tạp, xảy ra theo
mặt phẳng dọc của cơ thể. Ở các loài tảo của nhóm Desmokont cũng có sự phân
cắt dọc, mỗi tế bào con nhận một mảnh vỏ. Ở tảo Ceratium thì sự phân cắt xảy
ra theo mặt phẳng xéo và mỗi tế bào nhận một nửa vỏ phân chia theo một số
mảnh nhất định. Có khi một tế bào con nhận một roi.
Sinh sản hữu tính rất ít gặp ở tảo giáp. Ở một vài trường hợp sự sinh sản
này xảy ra giữa hai tế bào giống nhau.
Tảo giáp có vòng đời đơn bội (n), chỉ có nhân của hợp tử là đa bội (2n).
Tế bào thường tạo ra hợp tử nghỉ (hyponozygote). Chu trình sống: 31
Khoảng 50% số loài Dinophyta sống dị dưỡng (tế bào không có lục lạp),
một số lượng xấp xỉ như thế sống quang dưỡng, một số ít quang hợp nhưng lại thực bào.
Tảo giáp có một bào quan đặc biệt để bắt mồi, gọi là cuống (peduncle).
Đây là một cuống giả (pseudopod), có thể dãn dài ra, mọc ở vùng sulcus. Cuống
được sử dụng theo nhiều cách khác nhau tùy theo loài.
Các loài dị dưỡng ở nước biển của chi Protoperidinium thường sử dụng tảo Silic tàm thức ăn. Phân bố
Phần lớn tảo giáp sống ở biển (gần 90% tổng số loài) phần còn lại sống ở
nước ngọt. Khi tảo phát triển mạnh (còn gọi là nở hoa nước) sẽ làm thay đổi màu
của nước biển (có thể là màu đỏ, hoặc màu nâu, màu này là do sắc tố carotenoid
của tảo giáp). Hiện tượng này được gọi là thuỷ triều đỏ. Khi xảy ra hiện tượng
này sẽ làm ô nhiễm nguồn nước và ảnh hưởng đến các sinh vật khác sống trong
môi trường đó, bởi lẽ ngoài việc làm giảm lượng oxy hoà tan trong nước một số
các loài thuộc các chi như Alexandrium, Dinophysis, Prorocentrum.... còn có thể
tiết ra các độc tố gây tử vong không chỉ đối với sinh vật trong thuỷ vực mà còn
ảnh hưởng đến cả con người thông qua chuỗi thức ăn.
VII. Ngành tảo mắt - EUGLENOPHYTA
Ngành tảo này gồm chủ yếu là tảo đơn bào mang roi. Roi nẩy sinh từ đáy
của một huyệt gồm một rãnh và túi chứa.
Tế bào mang hai roi, roi ngắn nằm ở trong huyệt và roi dài mang một
hàng lông tơ mảnh gắn về một phía của roi cùng lớp với roi ngắn.
Lục lạp chứa chlorophyll a và b, không có chlorophyll c. Sắc tố phụ gồm:
carotein, neoxanthin, diatoxanthin, diadinoxanthin, zeaxanthin. 32
Một điểm mắt đỏ nằm tự do trong tế bào chất, sản phẩm dự trữ là
paramylon. Theo Gottlieb paramylon được cấu tạo bởi một carbonhydrat gần
giống với tinh bột nhưng không bắt màu với iodin, nên được gọi là các hạt
paramylon, chúng được xây dựng bởi kết nối (1,3 glucan là tinh thể có màng bao
(gồm hai phần: phần hình chữ nhật và phần có nhiều góc). Sản phẩm dự trữ dạng
lỏng - Chrysolaminarin có thể là một sản phẩm dự trữ thay thế ở một số loài tảo
mắt như Eutreptiella gymnasti, Sphenomonas leavis. Ở đây có thể gồm cả hai
loại. Paramylon hiện diện dưới dạng hạt có màng bao, chrysolaminarin ở trong
các túi ở phần cuối tế bào. Đa số tảo mắt sống ở nước ngọt, đặc biệt những thuỷ
vực giàu chất hữu cơ; một số ít loài ở biển. Ngành này có khoảng 40 chi, hơn
800 loài. Đa số loài có sắc tố, nhưng tảo mắt có khuynh hướng sống dị dưỡng.
Chi Euglena sống quang dưỡng nhưng cũng có quá trình dinh dưỡng các
hợp chất hữu cơ một cách mạnh mẽ. Đa số loài sống hoại sinh. Một số thực bào
(chi Peranema, Eutosiphon).
Tảo mắt sinh sản dinh dưỡng chủ yếu theo kiểu phân đôi tế bào. Tiến trình
phân đôi tế bào bắt đầu từ sự mở rộng về hai bên của huyệt và rồi từ đáy huyệt
một bộ roi mới được nảy sinh. Khi quá trình nguyên phân kết thúc, tế bào phân
dọc từ đỉnh đến cuối tế bào tạo nên hai tế bào con.
Sinh sản hữu tính chưa phát triển ở tảo mắt. Phân loại
Ngành có một lớp là Euglenophyceae, gồm 2 bộ Euglenales và Colaciales.
Một số chi thường gặp: 33
Chi Phacus: Tế bào đơn độc, bơi tự do trong nước, có hình dạng giống lá
trầu, không thay đổi, phẳng hay xoắn. Tế bào có hoặc không có điểm mắt. (Hình 2.31-1; 2.31-2).
Chi Euglena: Tế bào có một roi, luôn thay đổi hình dạng khi bơi trong
nước. Tế bào có dạng hình thoi đến hình kim với phần cuối tế bào hơi nhọn. Hầu
hết đều có điểm mắt ở phần đầu tế bào. (Hình 2.31-3 )
VIII. Ngành tảo lục - CHLOROPHYTA
Ngành tảo lục có khoảng 500 chi với 8.000 loài.
Ngành này bao gồm nhiều loài phiêu sinh đơn bào hoặc tập đoàn. Tuy
nhiên, cũng có nhiều dạng đa bào sống bám đáy, bám đá hoặc bám vào các giá
thể vào thời kỳ đầu của vòng đời nhưng sau đó tách rời giá thể sống trôi nổi tạo
thành những sợi tảo lớn.
Tảo lục chủ yếu sống ở nước ngọt, chỉ 10% sống ở biển. Các loài nước
ngọt thường phân bố có tính toàn cầu, chỉ một ít loài có tính đặc hữu cho vùng
sống. Ở môi trường biển, trong các vùng nhiệt đới thường có cấu trúc gần giống
nhau. Tuy nhiên, tính chất này không đúng với các loài sống ở vùng nước biển
lạnh: thành phần loài vùng Bắc và Nam bán cầu có sự khác biệt lớn. Một số tảo
lục sống khí sinh (aerophytic) trên vỏ cây, đất và đá, ngay trên cả tuyết và băng
cũng gặp tảo lục sinh sống, ví dụ loài Chlamydomonas nivalis gặp ở núi cao nơi
thường xuyên có tuyết. Cấu tạo:
Ở tảo lục, không có roi phủ hàng lông tơ vi ống 3 phần như ở
Heterokontophyta, các roi có cấu trúc giống nhau mặc dù có thể khác nhau về kích thước.
Thành tế bào bằng cellulose, tuy nhiên ở các bộ Caulerpales, xylans hay
manman thường thay thế cellulose.
Lục lạp bao quanh bởi một vỏ gồm hai lớp màng, không có lưới nội sinh
chất lục lạp bao quanh vỏ lục lạp. Thylakoid kết hợp thành dải gồm 3 - 4 cái.
Lục lạp chứa sắc tố chính giống với thực vật bậc cao, gồm chlorophyll a,
chlorophyll b và sắc tố lutein. Lục lạp có màu lục do sắc tố chlorophyll không bị
lấn át bởi các sắc tố phụ lutein, zeaxanthin, vioxanthin, antheraxanthin,
neoxanthin; đặc biệt các sắc tố siphonein, siphonixanthin gặp ở tế bào các chi tảo
dạng ống và chi tảo Tetraselmi. 34
Tảo lục có hai kiểu chuyển động: chuyển động hướng về ánh sáng (quang
hướng động) và tránh xa phía có ánh sáng (quang hướng nghịch). Tế bào mang
roi thường có điểm mắt. Điểm mắt luôn nằm trong lục lạp, phần trước tế bào và
gần gốc roi. Điểm mắt gồm một hay một số lớp các giọt lipid nằm trong phần cơ
chất giữa vỏ lục lạp và thylakoid ngoại vi tế bào. Sản phẩm dự trữ là tinh bột
nằm trong lục lạp thay vì ở trong tế bào chất, đây là đặc điểm khác với tảo có nhân thật còn lại.
Tảo lục sinh sản bằng các hình thức dinh dưỡng, vô tính và hữu tính, có các kiểu vòng đời sau:
+ Chu kỳ đơn kỳ đơn tướng:
Trong vòng đời chỉ có hợp tử là lưỡng bội (2n). ở một số tảo lục nước ngọt hợp
tử thường trải qua giai đoạn sống nghỉ - gọi là hợp tử ngủ (hypozygote), hợp tử
có vách dày sống nghỉ qua một thời gian mới nảy mầm.
+ Chu trình 2 kỳ đơn lưỡng tướng, có xen kẽ thế hệ đồng hình hay dị hình. Phân loại:
Lớp Micoromonadophyceace
Tế bào trần hay có vảy, thoi vô sắc ở vùng giữa còn tồn tại trong quá trình
phân bào (cytokinesis). Lớp gồm các tế bào tảo lục nguyên thủy mà một số tảo
trong đó được coi là nguồn gốc phát sinh của các lớp tảo lục khác của ngành. Lớp Prasinophyceae
Tảo lục chuyển động bằng roi, tế bào mang 1-8 roi gắn phía bên hay ở
đỉnh. Tế bào và roi phủ một số vảy chất hữu cơ.
Đại diện: Pyramimonas, Tetraselmis suecica. 35
Lớp Chlorophyceae
Đơn bào hay tập đoàn, chuyển động bằng roi. Một số dạng hạt, dạng
pamella, một số dạng sợi, tản và ống.Chủ yếu ở nước ngọt.
Lớp có khoảng 355 chi với 2.650 loài.
Tế bào sinh sản mang hai roi hoặc bốn roi ở đỉnh. Một số loài có tế bào
sinh sản mang roi, một số vừa hình thành tế bào sinh sản có roi vừa không roi.
Chu kỳ sống đơn kỳ, bào tử có giai đoạn nghỉ.
Bộ Volvocales: Đơn bào, tập đoàn chuyển động: 220 chi, 1.000 loài
Các đại diện của bộ: Chlamydomonas, Gonium (hình 2.34.1), Pandorina,
Eudogonium, Volvox (hình 2.34.2) Bộ Chlorococales
Chi Chlorococcum: Tế bào hình cầu hoặc elip có màng mỏng, đơn độc
hoặc liên kết với nhau lỏng lẻo. Thể màu hình chén hoặc hình lưới.
Chi Pediastrum: Cơ thể gồm 4-5-12 hoặc 16-3 -
2 64 tế bào phẳng liên kiết
với nhau giống hình bánh xe. Các tế bào ngoài cùng có từ 1-4 mấu với vết lõm
vào. Màng nhẵn hay gồ ghề, có nếp gấp.
Chi Scenedesmus: Gồm từ 2-4-8-1 -
6 32 tế bào liên kết với nhau bằng sườn
bên, xếp song song với nhau. Tế bào dài hình trụ hoặc oval. Các tế bào xếp
thành 1 hoặc 2 dãy hoặc xếp so le. Màng nhẵn hoặc phủ gai hay sừng.
Bộ Oedogoniales: đại diện Odogonium Lớp Ulvophyceae
Có khoảng 35 chi và 265 loài. Sống ở môi trường nước biển và lợ, chỉ một
số ít gặp ở nước ngọt, phổ biến là Ulothrix zonata. Ở sinh sản vô tính của 36
Ulothrix zonata, tất cả các tế bào có thể trở thành túi bào tử cho ra 16-32 động
bào tử gồm hai loại: loại nhỏ có 2 roi, loại to có 4 roi.
Khi sinh sản hữu tính, Ulva hình thành các giao tử hơi khác nhau. Túi giao
tử mọc thành cụm trên phiến.
Hợp tử sống chậm một thời gian và nẩy mầm không giảm nhiễm cho ra
cây bào tử lưỡng bội. Chu trình sống hai kỳ đơn lưỡng tướng và đồng hình luân phiên thế hệ.
Lớp Cladophorophyceae
Tản dạng hình ống, một số có cấu trúc phức tạp. Tế bào sinh sản mang 2-
4 roi. Mỗi tế bào của cây giao tử hoặc cây bào tử chứa nhiều nhân. Chu trình
sống gồm hai kỳ đơn lưỡng tướng đồng hoặc dị hình luân phiên thế hệ. Lớp Bryosidophyceae
Tảo luôn dạng ống, là một tế bào khổng lồ chứa nhiều nhân.
Tế bào sinh sản mang 2-4 roi. Chu kỳ sống 1 kỳ đơn tướng, trong đó cây
giao tử đơn bào và giai đoạn hợp tử lưỡng bội. Hợp tử có thể nẩy chồi thành cây
giao tử mới hoặc phân cắt cho ra động bào tử mang roi. Bộ Bryosidales:
Chu trình sống của Bryopsis plumosa đơn kỳ đơn tướng. Tảo dị tản
(dioecious) (heterothallie). Hợp tử có dạng sợi được cho là một loại bào tử chứa
một nhân lớn, có thể tạo nên động bào tử hoặc trực tiếp cho ra thể giao tử. Nếu
coi hợp tử dạng sợi là thể bào tử thì chu trình sống của chúng có sự luân phiên
thế hệ dị hình rất lớn.
Bộ Halimedales: Đại diện Chi Caulerpa 37
Tản gồm một phần bò và phần đứng, có nhiều hình dạng: dạng lá, dạng phiến xẻ thùy...
Một số tản đạt kích thước của một thực vật bậc cao dạng bụi. Tất cả
Caulerpa sống ở biển vùng nhiệt đới và cận nhiệt đới. Giao tử được tạo ra ở phần
đứng của tản và chúng được phóng thích qua các mấu lồi dạng một tên nhỏ.
Hợp tử sẽ phát triển thành một nguyên tản hình cầu (protosphere) chứa nhân hợp
tử. Sau đó nhiều sợi đa nhân được phát sinh từ nguyên cầu tản này và nảy nhánh
phát triển thành Caulerpa mới. Chu trình sống một kỳ.
Lớp Zygnematophyceae.
Gồm tảo dạng sợi và dạng hạt . Không nảy nhánh. Không có tế bào sinh
sản mang roi, sinh sản hữu tính theo kiểu tiếp hợp do hai giao tử dạng amip phối
hợp nhau, đặc điểm đặc trưng của lớp.
Tế bào có một nhân. Sắc thể có nhiều dạng, tế bào chứa 1 vài sắc thể hình dải xoắn.
Tảo lớp này có chu trình đơn bội. Trong vòng đời có qua một giai đoạn bào tử nghỉ (hypnospore) Bộ Zygnematales:
Tảo đơn nhân có các tế bào kết thành tập đoàn dạng sợi không nảy nhánh.
Bộ khoảng 18 chi với 900 loài ở nước ngọt, một số ít gặp ở đất đầm lầy.
Chi Spirogyra dạng sợi không nảy nhánh, chứa sắc thể hình dãi xoắn kiểu
lò xo. Sinh sản theo kiểu tiếp hợp. Chu trình sống đơn kỳ đơn tướng.
Chi Zygnema chứa 2 sắc thể hình sao nằm theo trục tế bào, và một cái
chứa hạt tạo bột. Bộ Desmidiales
Tảo phần lớn đơn bào, nhưng một số có dạng tập đoàn không ổn định hình
dạng hoặc sợi không nẩy nhánh. Thành tế bào gồm hai nửa đối xứng nối nhau ở
1vùng eo hẹp. Thành tế bào có nhiều chạm trổ khác nhau. Trên thành tế bào 38
cũng có các lỗ hoặc những cấu trúc biến đổi dạng lỗ mà qua đó chất nhầy thường
được tiết ra. Bộ có khoảng 30 chi với 500 loài. Tảo của bộ này đa dạng và hình
dáng đẹp, thường phát triển ở vùng nước ngọt có pH thấp.
Các đại diện tiêu biểu: Gonatozygon, Closterium, Cosmarium, Euastrum,
Micrasterias, Staurastrum...
Chi Micrasterias: Tản đơn bào. Kích thước lớn, chiều dài lớn hơn chiều
rộng. Eo thắt ở giữa luôn sâu và gần như thẳng. Đường bao ngoài của nửa tế bào
khi nhìn ở mặt trước có dạng bán nguyệt đến hình lục giác. Nửa tế bào thường
có các đường rạch tạo thành các cạnh. Tế bào có thành mềm và một số có lớp gai nhỏ bên ngoài.
Chi Cosmarium: Tế bào eo thắt sâu ở giữa, chiều dài hơi lớn hơn chiều
rộng. Thành tế bào mềm hoặc có các vân lỗ hoặc có các u sần kiểu mụn cóc,
không có gai. Mặt trước của nửa tế bào hình bán cần, elip, hình thận, hình thang hoặc gần vuông.
Chi Closterium: Tế bào dài, cong hoặc có hình bán nguyệt rõ ràng, thon
dần về hai đầu, không có eo thắt ở giữa. Thành tế bào có các lỗ nhỏ. Tế bào màu vàng nâu hoặc nâu.
Lớp Charophyceae
Tảo có cơ thể lớn, có thể cao tới vài cm. Tản dạng sợi và dạng ống, phân
thành mấu và lóng. Mỗi mấu mang một vòng nhánh, có sinh trưởng giới hạn.
Các tế bào trưởng thành chứa nhiều nhân, tế bào lúc non đơn nhân.
Tất cả tảo Charophyceae sinh sản hữu tính noãn giao. Túi trứng luôn được
mang ở nhánh bên gồm trứng bao quanh bới các tế bào hẹp. Túi tinh cũng được
mang trên một nhánh và chứa nhiều sợi sinh giao tử đa bào. Mỗi tế bào của sợi
sẽ cho một giao tử. Xung quanh túi giao tử là một vỏ bao gồm các tế bào áo bất thụ.
Lớp này chủ yếu phân bố ở nước ngọt, một số ít ở nước lợ.
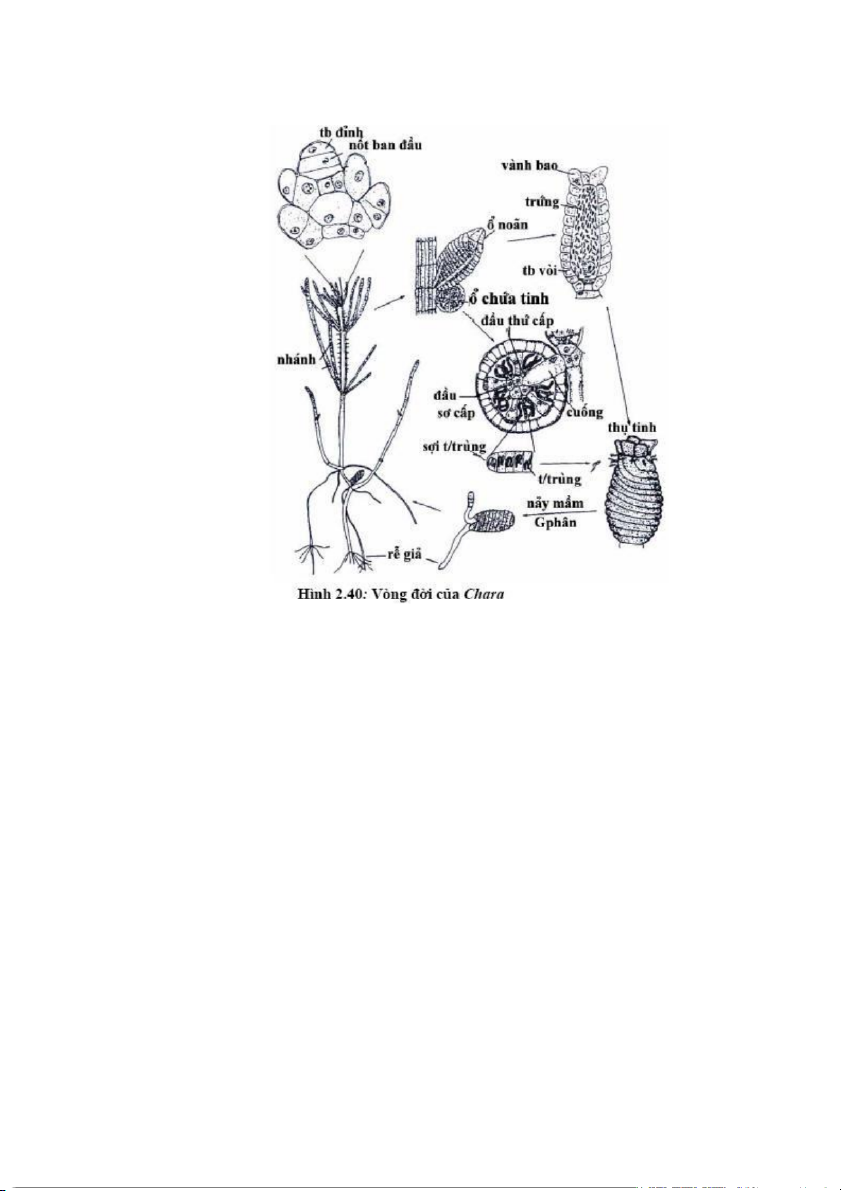
Các đại diện tiêu biểu: Chara, Nitella, Laprothamnium Chi Chara
Tế bào lóng của Chara dài và lớn, được bao quanh bởi những dãy tế bào
kéo dài tạo nên một lớp vỏ. Ở mỗi mấu mang một vòng “lá”. Các lá của Chara
cũng phân thành mấu và lóng nhỏ ở trục chính. Ở đây mỗi mấu nảy sinh một
vòng gai đơn bào, đôi khi được gọi là "lá nhỏ". Ở các loài đồng tản, cơ quan sinh 39
giao tử đực hình cầu và cơ quan sinh sản cái hình độc bình thường nằm từng cặp
ở nách “lá”. Cây bám vào bùn bằng rể giả. Chi này có khoảng 20 loài.
Cấu trúc cơ quan sinh sản đực và cái rất phức tạp. Cả hai cơ quan này
được phát sinh từ tế bào bên của mấu lá. Tế bào đầu tiên phân cắt từ tế bào khởi
sinh túi tinh là tế bào cuống. Tế bào ngoài cùng phân cắt tạo ra 4 tế bào rồi 8 tế bào.
Mỗi tế bào này phân cắt tạo ra một tế bào ở ngoài gọi là tế bào áo, tế bào ở
giữa gọi là tế bào chống (manubrium) tiếp đến là tế bào đầu (capitulum) sơ cấp.
Tế bào phát triển lớn ra về mặt bên và các tế bào manubrium và capitulum được
kéo ra tạo nên một khoảng trống trong cơ quan sinh giao tử. Tế bào manubrium
kéo dài xuyên tâm trong khi tế bào capitulum tiếp tục phân chia tạo nên 6 tế bào
capitulum thứ cấp. Từ tế bào capitulum này sẽ sinh ra các sợi sinh tinh gồm
khoảng 5-50 tế bào, mỗi tế bào sinh tinh sẽ sản sinh một giao tử chuyển động.
Tế bào sinh cơ quan sinh sản cái phân chia cho ra 3 tế bào. Tế bào trên
cùng phát triển thành tế bào trứng, tế bào phía dưới phát triển cho ra tế bào bao
quanh trứng, tế bào thứ 3 phát triển thành cuống tế bào trứng. 40
CHƯƠNG 4. RONG BIỂN
Rong biển là những loại tảo, thuộc thực vật bậc thấp, phân bố ở các vùng
cửa sông, vùng nước lợ, vùng triều cho đến các vùng biển sâu, có cấu tạo đơn
bào hoặc đa bào, cơ thể chưa phân thành rễ, thân, lá thật nên gọi là tản (Thallus);
toàn bộ cơ thể có chung một chức năng tự dưỡng, quá trình phát sinh không có
giai đoạn phôi thai mà chỉ có hợp tử, hợp tử có khả năng phát triển độc lập với cơ thể mẹ.
I. MỘT SỐ ĐẶC ĐIỂM SINH HỌC CỦA RONG BIỂN
1. Quá trình phân cắt và sự phát sinh của rong biển
- Hình thành khối đa bào
Dù bào tử bám có cực hoặc không có cực, phân cắt lần 1 là phân cắt dọc
và cho ra 2 tế bào. Phân cắt lần 2 có thể phân cắt dọc hoạc ngang (tùy loài), hình
thành 4 tế bào. Những lần phân cắt tiếp theo không tuân theo một quy tắc nhất
định, hình thành khối đa bào cân đối (có cực) hoặc không cân đối (không có cực).
- Sự phát triển giai đoạn
Từ khối đa bào trở đi, các giai đoạn khác nhau của cơ thể được tiếp tục
phát triển gọi là phát triển giai đoạn. Do khối lượng tế bào và hình dạng khối đa
bào thay đổi nên sự phát sinh khác nhau, làm cho hình thái cấu tạo của cơ thể
trưởng thành cũng khác nhau. Nếu cơ thể trưởng thành có cơ quan bám là hình
đĩa giác bám thì dạng đĩa của khối đa bào chính là cơ quan bám của cơ thể
trưởng thành. Dạng phát sinh này thấy ở các giống Gracilaria, Hypnea... Nếu cơ
thể trưởng thành có cơ quan bám là rễ giả thì khi phát sinh, khối đa bào sẽ phát
triển theo hướng xuống sát vật bám, một số tế bào kéo dài họp lại thành rễ giả
phân nhánh. Dạng này có ở Gelidium, Asparogopsis...
Sau khi hình thành cơ quan bám, ở trung tâm khối đa bào xuất hiện tế bào
phân sinh (tế bào mầm). Tế bào mầm phát triển thành mầm, cây mầm và cuối
cùng là cây trưởng thành.
Rong biển bậc cao: có 2 dạng hình thái và cấu tạo tùy thuộc vào 2 hình
thức phát sinh. Đó là dạng phân nhánh có cấu tạo đa trụ và dạng đơn nhánh có cấu tạo đơn trụ.
- Dạng đơn trụ: tế bào mầm phân cắt lần 1 là phân cắt chéo 450 so với trục
đỉnh, cho ra 2 tế bào (TB’ (trên) và TB” (dưới)). Tế bào TB’ phân cắt lần 2, lần
3 là phân cắt dọc cho ra 4 tế bào sau này thành 4 hàng tế bào vây trụ. Tế bào
TB” chỉ phân cắt ngang, hình thành 1 hàng tế bào trung trụ.
- Dạng đa trụ: quá trình phân cắt lần 1 như trên. Lần phân cắt thứ 2 và 3,
cả tế bào TB’ và TB” đếu phân cắt dọc. TB’ hình thành những hàng tế bào vây
trụ. TB” hình thành những hàng tế bào trung trụ. Tế bào vây trụ phân cắt dọc,
ngang hình thành những hàng tế bào nội bì và ngoại bì.
Quá trình phân nhánh: tương tự quá trình hình thành thân. Tế bào mầm
được hình thành do một trong các tế bào ngoài cùng của tầng lõi (gồm tế bào 41
trung trụ và vây trụ) sau khi phát triển ra mặt ngoài của cơ thể. Phân cắt lần 1 là
phân cắt chéo như tế bào mầm của khối đa bào đầu tiên ở dạng đĩa. Nếu thân
chính có cấu tạo đơn trụ thì nhánh có cấu tạo đơn trụ. Nếu thân chính có cấu tạo
đa trụ thì nhánh có cấu tạo đa trụ.
Rong hình bản, hình lá: sau khi hình thành khối đa bào và cơ quan bám,
tất cả các tế bào trong khối đa bào dạng đĩa đều phân cắt không quy tắc theo
nhiều hướng khác nhau tạo thành dạng hình bản, hình lá, hình ống... Khi phát
sinh theo dạng này, dạng đĩa của khối đa bào không hình thành tế bào mầm.
Rong hình sợi: là dạng có tế bào mầm nhưng tế bào này chỉ phân cắt
ngang, do đó các tế bào con chỉ phát triển theo một hướng nhất định, kéo thành
sợi. Vì vậy, hình sợi thực chất do 1 hàng tế bào tạo nên. Chỗ phân nhánh trên sợi
là do một tế bào phát triển mạnh hơn tế bào khác rồi phân cắt mà thành.
2. Đặc điểm phân chia giai đoạn phát triển của rong biển
Sự phân chia giai đoạn của rong biển trong sinh học
Toàn bộ chu kỳ sinh sản của một loài rong được chia thành 5 giai đoạn
như sau: phát sinh, sinh trưởng, tích lũy, sinh sản, tàn lụi.
Sự phân chia giai đoạn phát triển của rong biển trong sản xuất
Giai đoạn cây mầm: là giai đoạn sinh bào tử, phân cắt phát triển thành
mầm rồi cây mầm. Đây là khâu đầu tiên trong chu kỳ sản xuất. Tỷ lệ thành mầm
của bào tử rất thấp nhưng khi đã thành mầm nó sẽ tăng trưởng nhanh chóng để
chuyển sang giai đoạn cây giống. Thời gian của giai đoạn này phụ thuộc vào thời
kỳ phát sinh của mỗi loài rong biển. Phần lớn các loài rong có mầm phát sinh từ
bào tử có kích thước bé hơn cây mầm phát sinh từ sinh sản sinh dưỡng. Thời kỳ
phát sinh của mầm cũng dài hơn. Ví dụ cây mầm của rong Câu chỉ vàng
Gracilaria asiatica kích thước 1cm phát sinh từ bào tử mất 40-45 ngày nhưng
nếu phát sinh từ mầm dinh dưỡng chỉ mất 18-20 ngày.
Giai đoạn cây giống: là thời kỳ tiếp theo của giai đoạn cây mầm, ứng với
giai đoạn sinh trưởng. Kích thước của cơ thể ở giai đoạn này thường căn cứ vào
thời gian sinh trưởng, (được gọi là các đơn vị thời gian sinh trưởng: ngày tuổi,
tháng tuổi) và phụ thuộc nhiều vào kích thước cây rong ở giai đoạn trưởng
thành. Về hình dạng, ở giai đoạn này, những loài có phân nhánh phần lớn chưa
xuất hiện nhánh cấp 2, 3 tương đương với giai đoạn sinh trưởng. Về mặt các
chức năng sinh lý, rong ở giai đoạn này có khả năng thích ứng cao với điều kiện môi trường.
Giai đoạn cây trưởng thành: về hình thái, cây trưởng thành phát triển hoàn
chỉnh các bộ phận trên cơ thể. Những loài phân nhánh đã có đầy đủ nhánh các
cấp. Ở thời kỳ sau, trên cơ thể đã xuất hiện các cơ quan sinh sản, màu sắc cơ thể
rong bóng bẩy, đậm đà, rong bụ bẫm, nhánh vươn dài, sự tích lũy chất dinh
dưỡng tăng lên. Lúc này hiện tượng tàn lụi cũng xuất hiện rải rác ở đầu nhánh.
Giai đoạn cây trưởng thành ứng với giai đoạn cuối sinh trưởng, tích lũy, sinh sản
và một phần giai đaọn tàn lụi. Đây là giai đoạn cây rong đang trong thời gian
nuôi trồng hoặc là giai đoạn cây sản phẩm, là thời kỳ cuối trong chu kỳ sản xuất. 42
Việc xác định đúng giai đoạn cây trưởng thành có ý nghĩa quan trọng trong kế
hoạch sản xuất (chăm sóc, thu hoạch, lịch sản xuất cho vụ sau).
3. Đặc điểm sinh thái của bào tử - Sự phát tán
Phát tán là quá trình bào tử thoát khỏi cơ thể mẹ ra môi trường nước. Đó
là biểu hiện của thời kỳ hậu sinh sản. Sự phát tán của giống tuân theo các quy luật:
+ Tháng phát tán: mỗi năm chỉ có một tháng mà trong tháng đó bào tử
được phóng ra nhiều nhất. Quy luật này đúng với rong sống nhiều năm, có quy
luật sinh trưởng, sinh sản nhất định ở các tháng trong năm.
+ Con nước phát tán: trong ngày triều cường, nước rút xuống ở mức thấp
nhất, thời gian cơ quan sinh sản của rong biển lộ ra ngoài không khí dài nên số
lượng cá thể rong biển được kích thích khô tăng lên... Khi nước lên cao, xuống
mạnh càng làm tăng nhân tố chấn động do vậy bào tử phóng ra càng nhiều. Quy
luật này đúng với rong phân bố ở vùng triều.
+ Giờ phát tán: vào ngày con nước cường, giờ phát tán trùng với giờ cao
của thủy triều. Ngày thường, giờ phát tán trùng với giờ cao điểm của tổng hợp
các yếu tố nhiệt độ, độ mặn, chấn động... - Sự di động
Sau khi phát tán ra khỏi cơ thể mẹ, bào tử di động một thời gian trước khi
bám vào vật bám. Các loại bào tử khác nhau có sự di động khác nhau. Bảo tử
động di động chủ động nhờ tiên mao. Bào tử bất động di động thụ động nhờ tác
dụng của dòng nước hoặc chuyển động biến hình. Thời gian di động của bào tử
phụ thuộc các yếu tố ngoại cảnh, đặc biệt là nhiệt độ. Ngoài ra, còn phụ thuộc
vào sự chín muồi của bào tử. Ví dụ thời gian di động của bảo tử được phóng
thích nhân tạo ngắn hơn so với bào tử được phóng thích tự nhiên. - Quá trình bám
Sau khi di động, bào tử bám vào vật bám để phân cắt và phát triển. Các
loại bào tử khác nhau có phương thức bám khác nhau.
II. ẢNH HƯỞNG CỦA CÁC YẾU TỐ MÔI TRƯỜNG ĐẾN SỰ
SINH TRƯỞNG - PHÁT TRIỂN CỦA RONG BIỂN
1. Yếu tố động lực a. Thủy triều
Thủy triều là hiện tượng lên xuống của nước biển có quy luật, dưới tác
dụng của lực hấp dẫn giữa mặt trăng, mặt trời, trái đất và các thiên thể khác.
Giới hạn nước lên cao nhất và rút thấp nhất gọi là vùng triều. Thủy triều có ảnh
hưởng đến sự phân bố của rong biển. Rong biển hầu hết tập trung phân bố từg
vùng trung triều trở xuống. Ở vùng trên triều, rong ít phân bố, thường chỉ có các
loài rong có kích thước nhỏ, có tính chịu khô cao, đại diện nhóm này có giống 43
Bostrychia thuộc ngành rong Đỏ. Vùng trung triều và hạ triều, nhất là tầng trên
của vùng dưới triều thường tập trung các loại rong có kích thước lớn.
Sự lên xuống của thủy triều còn tác động đến quá trình phóng bào tử của
rong biển. Rong Mơ Sargassum, rong Quạt Padina có chu kỳ phóng bào tử phụ
thuộc vào chu kỳ thủy triều. Rong câu chỉ vàng Gracilaria asiatica phân bố vùng
triều, khi nước triều rút, rong bị khô có tác dụng kích thích bào tử quả phóng khi nước triều dâng lên. b. Sóng gió
Những loài rong ưa sóng gió thường phân bố ở các vùng có sóng lớn,
thường ở vùng triều. Đại diện cho nhóm này là Gelidium amansii, Hypnea,
Ecklonia, Chaetomorpha… Nhóm rong này có cơ quan bám phát triển, cấu tạo cơ thể rắn chắc.
Những loài rong sợ sóng gió thường phân bố ở vùng sóng yếu hoặc ở
vùng nước tĩnh như trong các ao đầm nước lợ. Đại diện cho nhóm này là
Gracilaria, Enteromorpha, Ulva, Monostroma… Nhóm rong này có cơ thể mềm
mại, cấu tạo trong lỏng lẻo.
Quá trình bám của bào tử rong biển phụ thuộc vào sóng gió. Những loài
rong sống ở vùng sóng lớn, bào tử của nó phóng ra thường có tốc độ bám nhanh
hơn những loài phân bố ở vùng yên sóng. Sóng gió còn là yếu tố có tác dụng cơ
học cho quá trình phóng bào tử của rong biển. c. Hải lưu
Hải lưu là sự di chuyển của dòng nước biển có quy luật, có liên quan đến
sự thay đổi nhiệt độ của vùng nước. Hải lưu có ảnh hưởng lớn đến sự di động
phát tán của bào tử rong biển, ảnh hưởng đến hoạt động dinh dưỡng và phân bố của rong biển. 2. Yếu tố vật lý
a. Địa bàn sinh trưởng của rong biển
Căn cứ vào tập tính sống, có thể chia rong biển thành 2 dạng: sống cố định
và sống phù du. Những loài tảo sống phù du thường phân bố ở các tầng nước
khác nhau, đảm bảo cho chúng có khả năng hấp thụ năng lượng ánh sáng mặt
trời để quang hợp. Chúng thường không có cơ quan bám. Đối với rong biển sống
cố định thì trong quá trình sống chúng cần có địa bàn sinh trưởng. Địa bàn sinh
trưởng của rong biển có thể là đáy cứng như đá tảng, đá cuội, san hô… hoặc đáy
mềm như bùn, bùn cát, cát bùn hoặc cơ thể thực vật khác cùng phân bố với
chúng. Rong biển hấp thụ nước, muối khoáng từ môi trường xung quanh chứ
không phải từ địa bàn sinh trưởng. Địa bàn sinh trưởng chỉ giúp chúng cố định ở
một tầng nước nhất định trong quá trình sống để đảm bảo thực hiện tốt quá trình quang hợp.
Địa bàn sinh trưởng có quan hệ đến quá trình hình thành cơ quan bám và
khả năng bám của rong biển. Yêu cầu về dạng địa bàn sinh trưởng ở rong biển
khác nhau tùy loài. Rong sống ở vùng triều có cơ quan bám phát triển, thích bám 44
trên các dạng đáy cứng. Chẳng hạn như rong Thạch Gelidium thích phân bố trên
tảng đá có nhiều động vật thân mềm, rong Chuỗi Chaetomorpha thích bám trên
đáy có nhiều cát sỏi… Rong sống trong đầm nước lợ, cơ quan bám kém phát
triển, chúng thường sống tự do cài quấn hoặc một phần gốc vùi trong lớp bùn
cát, một số loài sống bám trên thực vật thủy sinh khác. Chẳng hạn như rong
Guột Caulerpa có rễ giả phát triển đâm sâu trong lòng cát, rong Đen đầu
Sphacellaria thường bám trên gốc rong Mơ; rong Nhiều ống Polysiphonia
thường bám trên rong Câu. b. Nhiệt độ
Nhiệt độ ảnh hưởng đến sự phân bố của rong biển. Căn cứ vào quan hệ
giữa nhiệt độ và phân bố của rong biển, Kjellman và Sberkoj đã phân chia khu
hệ rong biển theo thanh nhiệt độ như sau:
Nhiệt độ 0-50C ứng với khu hệ rong hàn đới. Nhiệt độ 5-15 C 0
ứng với khu hệ rong á hàn đới
Nhiệt độ 10-200C ứng với khu hệ rong ôn đới
Nhiệt độ 15-250C ứng với khu hệ rong á nhiệt đới
Nhiệt độ ≥ 250C ứng với khu hệ rong nhiệt đới
Dựa theo cách phân chia này thì khu hệ rong biển của Việt Nam mang
tính chất á nhiệt đới và nhiệt đới từ bắc vào nam. Đặc điểm của rong biển ở các
khu hệ đó có sự khác nhau rõ ràng. Rong trong khu hệ rong hàn đới và ôn đới có
kích thước cơ thể tương đối lớn, thời gian sinh trưởng muộn và kéo dài, số lượng
cá thể trong cùng một loài nhiều nhưng thành phần loài kém đa dạng. Rong
thuộc khu hệ rong nhiệt đới thì có đặc điểm ngược lại.
Trong phạm vi nhiệt độ thích hợp, thì quá trình sinh trưởng của rong biển
tăng và ngược lại. Yêu cầu nhiệt độ thích hợp cho quá trình sinh trưởng của rong
biển khác nhau tùy loài. Rong Mứt Porphyra tenera sinh trưởng tốt ở nhiệt độ
trên dưới 100C; rong Quạt Padina sinh trưởng tốt ở nhiệt độ 20-250C, rong Unda
Undaria pinnatifera sinh trưởng tốt ở nhiệt độ 200C .
Nhiệt độ ảnh hưởng đến quá trình sinh lý của rong biển. Quá trình quang
hợp và hô hấp của rong biển được tiến hành thuận lợi trong phạm vi nhiệt độ
thích hợp. Rong Câu chỉ vàng Gracilaria asiatica có cường độ quang hợp lớn
nhất trong giới hạn nhiệt độ 25-280C. Khi nhiệt độ nước tăng lên quá 350C hoặc
nhỏ hơn 100C thì quá trình quang hợp của rong bị giảm nhanh chóng.
Nhiệt độ ảnh hưởng đến chức năng sinh sản của rong biển. Yếu tố nhiệt
độ thích hợp thúc đẩy quá trình sinh trưởng của rong biển và khi đạt đến giai
đoạn sinh trưởng – phát triển nhất định, rong sẽ thực hiện quá trình sinh sản.
Mùa vụ sinh sản của rong biển phụ thuộc vào nhiệt độ môi trường. Ở nước ta,
mùa vụ sinh sản của rong biển xảy ra vào khoảng tháng 3-5 khi mà nhiệt độ
nước thích hợp cho quá trình hình thành cơ quan sinh sản ở rong biển. Mỗi loài
rong khác nhau yêu cầu nhiệt độ thích hợp cho quá trình sinh sản có sự khác
nhau. Rong Quạt Padina là rong á nhiệt đới, ở nhiệt độ 20-250C mới có thể phát 45
dục. Rong Câu chỉ vàng Gracilaria asiatica hình thành túi bào tử quả rộ trong
thời gian tháng 4-5, và bào tử quả phóng tốt trong điều kiện nhiệt độ 250C . c. Ánh sáng
Năng lượng mặt trời chiếu xuống trái đất rất lớn nhưng thực vật trong quá
trình quang hợp chỉ đồng hóa 1-3% năng lượng mặt trời chiếu trên lá. Đối với
thực vật thủy sinh, năng lượng đồng hóa được còn nhỏ hơn con số trên. Những
tia bức xạ mặt trời thấy được có độ dài sóng trong khoảng 380-780nm. Trong
các tia nhìn thấy này, tia đỏ (600-780nm) có tác dụng rất lớn trong quang hợp.
Tia hồng ngoại có độ dài sóng lớn nhất (780-340.000nm), mắt thường không
thấy được, tia này không có tác dụng sinh trưởng đối với thực vật mà chỉ có ảnh
hưởng gián tiếp đến thực vật qua tác dụng nhiệt. Ánh sáng chiếu xuống thủy vực
khuếch tán thành các phần ánh sáng tán xạ, ánh sáng phản xạ và ánh sáng hấp
thụ. Cường độ ánh sáng tán xạ yếu nên thuận lợi cho sự phát triển của thực vật.
Theo L.A Ivanop, ánh sáng tán xạ có 50-60% tia sinh lý (tia có tác dụng tới
quang hợp), còn ánh sáng phản xạ và ánh sáng hấp thụ ít có tác dụng cho sự phát triển của thực vật.
Các vùng sáng trong thủy vực được phân ra như sau:
Từ 0-200m: vùng sáng, là vùng có đủ các tia sáng.
Từ 200-1500m: vùng mặt sáng, là vùng có các tia sóng ngắn, cực ngắn,
chủ yếu là ánh sáng tím.
Từ trên 1500m: vùng tối, vùng không còn tia sáng nào xâm nhập tới.
Ở các thủy vực nước ngọt, độ trong thấp hơn nên giới hạn các vùng này
nhỏ hơn ở biển. Vùng sáng khoảng vài chục mét , vùng mặt sáng từ vài chục đến 200m, vùng tối trên 200m.
Ánh sáng ảnh hưởng đến sự phân bố của rong biển theo chiều thẳng đứng.
Rong Xanh Chlorophyta có nhiều diệp lục tố thích hợp hấp thụ các tia sáng đỏ,
thường phân bố ở tầng mặt có độ sâu 5-6m. Rong Nâu Phaeophyta có nhiều sắc
tố phụ (phycophein và fucoxanthyl) thích ánh sáng da cam, vàng nên thường
phân bố ở tầng giữa, sâu khoảng 30-60m. Rong Đỏ Rhodophyta có nhiều sắc tố
phụ Phycoerythrin và Phycoxianin thích ánh sáng xanh nên thường sống ở tầng
nước sâu nhất khoảng 100m. Rong Mứt Porphyra, rong Cải biển Ulva, rong Bún
Enteromorpha ưa ánh sáng mạnh nên thường phân bố ở vùng cao và trung triều.
Những loài rong này thích ánh sáng mạnh như thực vật trên cạn nên gọi là thực
vật dương tính. Rong Unda Undaria pinnatifera, rong Bẹ Laminaria japnica
thích vùng ánh sáng yếu nên thường sống ở độ sâu 3-5m dưới tuyến hạ triều,
thuộc nhóm thực vật ưa ánh sáng yếu giống như thực vật âm tính lục địa.
Ánh sáng ảnh hưởng đến sinh trưởng, sinh sản của rong biển. Ở Việt
Nam, vụ Đông xuân có cường độ ánh sáng thích hợp cho quá trình sinh trưởng
của rong biển. Mỗi loài rong khác nhau, yêu cầu ánh sáng cho quá trình quang
hợp cũng khác nhau. Rong Câu chỉ vàng Gracilaria asiatica có cường độ quang
hợp lớn nhất khi cường độ ánh sáng đạt 40.000-50.000lux. Rong Mứt Porphyra
quang hợp mạnh trong giới hạn ánh sáng khoảng 100-1000lux. Rong Unda 46
Undaria pinnatifera, rong Bẹ Laminaria japnica quang hợp tốt trong giới hạn
ánh sáng khoảng 1000lux. Ánh sáng còn ảnh hưởng đến quá trình nảy mầm của
rong biển. Đối với rong Mứt Porphyra tenera khi cường độ ánh sáng trong
khoảng 100-1200lux thì thể sợi sinh trưởng tốt, nếu ánh sáng giảm xuống 6-
16lux, màu sắc rong nhợt nhạt, thể sợi bị teo dần. 3. Yếu tố hóa học a. Độ mặn
Độ mặn có ảnh hưởng đến sự phân bố của rong biển. Khoảng 90% giống
loài rong Đỏ, rong Nâu phân bố ở biển trong khi chỉ có khoảng 10% giống loài
rong Xanh phân bố ở nước mặn, lợ. Dựa vào khả năng thích nghi với độ mặn của
môi trường, người ta phân chia rong biển thành các nhóm sau:
Nhóm hẹp muối ở độ mặn cao: bao gồm những loài rong đặc trưng của
vùng triều và biển sâu, chúng phân bố và sinh trưởng ở những nơi có độ mặn cao khoảng 25-36‰.
Nhóm hẹp muối ở độ mặn thấp: gồm những loài rong chỉ xuất hiện trong
các đầm nước lợ vào mùa mưa, khi độ mặn môi trường thấp.
Nhóm rộng muối: gồm những loài rong có khả năng phân bố từ vùng triều
đến các ao đầm nước lợ.
Độ mặn có ảnh hưởng đến quá trình sinh trưởng của rong biển. Yêu cầu
độ mặn thích hợp cho quá trình sinh trưởng của mỗi loài rong có sự khác nhau.
Rong Bẹ Laminaria japnica sống ở độ mặn 30-31‰, khi độ mặn giảm xuống
28‰ thì rong sinh trưởng kém. Rong Mứt Porphyra tenera chịu sự biến động độ
mặn tương đối lớn, chúng sống được ở độ mặn 26-32‰, khi độ mặn giảm xuống
dưới 26‰ rong vẫn có thể sống được. Rong Câu chỉ vàng Gracilaria asiatica
thuộc loài rộng muối, có thể sống được ở vùng triều đến các ao đầm nước lợ,
sống trong giới hạn độ mặn 5-30‰, có khả năng quang hợp tốt ở độ mặn 20- 25‰.
III. ĐẶC ĐIỂM SINH THÁI CỦA KHU HỆ RONG BIỂN VIỆT NAM 1. Miền Bắc
Vùng biển ven bờ có 2 loại hình môi trường có rong biển phân bố: vùng
triều và vùng nước lợ.
Vùng triều: là vùng ven biển, ven đảo có độ mặn cao, sóng đánh mạnh,
nước trong và chịu tác động thường xuyên của chế độ thủy triều. Hầu hết rong
biển tập trung phân bố ở vùng này. Chất đáy mềm (bùn cát) thường hình thành
vùng gần cửa sông, trong những vịnh kín hoặc nửa kín. Loại chất đáy này thích
hợp với những loài rong có rễ giả hoặc những loài ưa sống cài quấn. Đại diện
tiêu biểu là Caulerpa, Enteromorpha, Cladophora, Gracilaria. Chất đáy cứng phổ
biến ở các vùng ven đảo, các mũi bờ biển nơi có đá tảng hoặc nơi kéo dài của
chân núi đá san hô, đá cuội trên nền cát, chỗ sóng đánh mạnh. Đại diện của
những loài ưa chất đáy cứng là Gelidium, Hypnea, Gracilaria. 47
Vùng nước lợ: được hình thành do việc đắp đê khoanh vùng để nuôi trồng
thủy sản. Đặc điểm vùng này là luôn có nước, đáy bùn cát, có độ mặn biến đổi
theo mùa, ít bị xáo trộn bởi các tác nhân vật lý của biển. Rong biển trong vùng
này nghèo về thành phần loài, chỉ những loài có nguồn gốc ở biển xâm nhập vào
có biên độ sinh thái rộng mới phát triển với số lượng cá thể lớn. Các đại diện
điển hình là Enteromorpha, Chaetomorpha, Lyngbya, Gracilaria. 2. Miền Nam
Có số lượng loài phong phú hơn, tập trung ở vùng ven biển miền Trung,
ven các đảo. Phần lớn các loài ở đây thích nghi với chất đáy cứng hoặc sống trong đầm nước lợ.
3. Tính chất khu hệ rong biển Việt Nam
So với khu hệ rong biển nhiệt đới chính thức thì khu hệ rong biển phía bắc
có ít loài rong nhiệt đới nhưng có nhiều loài rong cận nhiệt đới. Vì vậy, khu hệ
rong biển miền Bắc Việt Nam thuộc vào vùng nhiệt đới của khu phụ cận nhiệt
đới Bắc bán cầu. Điều này có thể được giải thích bởi bờ biển Bắc Việt Nam chịu
ảnh hưởng của nhiều dòng nước lạnh chảy từ phía Bắc vào vịnh qua eo biển Hải Nam.
Tính chất khu hệ rong biển phía Nam: biển phía Nam chịu ảnh hưởng trực
tiếp của dòng nước nóng nhiệt đới, vì vậy số loài rong nhiệt đới chiếm tỷ lệ cao,
những loài rong cận nhiệt ít. Như vậy, khu hệ rong biển phía Nam mang tính
chất nhiệt đới điển hình và gần với khu phụ nhiệt đới thật sự.
Tính chất khu hệ rong biển miền Bắc Việt Nam rất phức tạp, được chia thành 2 vùng rõ rệt:
Vùng 1: Quảng Ninh đến bắc Hải Phòng
Do tính chất địa lý bờ biển phức tạp, khúc khuỷu, có nhiều đảo nhỏ, độ
chênh lệch thủy triều rất lớn (3,8-6,7m), độ mặn vùng cửa sông dao động từ 9,3-
13‰, độ mặn ở các đảo là 30-32‰, nhiệt độ nước mùa đông là 14-17 C, 0 mùa hè
khoảng 29,70C, rong biển vùng này phát triển tập trung, số lượng và kích thước cá thể lớn.
Vùng 2: Từ nam Hải Phòng trở vào
Bờ biển bằng phẳng, ít khúc khuỷu, vùng này có điều kiện tự nhiên gần
với vùng đầm hồ và vùng biển nhiệt đới. Thành phần loài rong phong phú, rong có kích thước nhỏ.
IV. MÙA SINH HỌC CỦA RONG BIỂN
Hầu hết những loài rong biển sống ở vùng triều ven biển Việt Nam có
mùa tương đối rõ rệt, chu kỳ sinh trưởng 1 năm, phát triển mạnh vào các tháng
3-5. Những loài sinh trưởng nhiều năm có thời gian tồn tại lâu hơn, hầu như
quanh năm, nhất là những loài sống ngập ở bãi triều. Dựa vào đặc điểm phát
triển, người ta chia rong biển thành các nhóm loài sau:
- Nhóm rong chỉ phát triển vào mùa lạnh hoặc chỉ kéo dài đến những
tháng chuyển tiếp đầu mùa nắng như Porphyra, một số loài trong giống 48
Chaetomorpha, Enteromorpha. Phần lớn những loài rong nhóm này mang tính
chất của rong vùng nước lạnh.
- Nhóm rong phát triển cuối mùa lạnh sang cả mùa nóng. Bao gồm những
loài rong mang tính chất rong nước nóng, phát triển quanh năm. Nhóm này
chiếm ưu thế ở vùng vịnh Bắc bộ. Tuy nhiên do chế độ nhiệt của không khí và
nước ở phía bắc và phía nam vịnh Bắc bộ khác nhau, nhất là mùa lạnh nên mùa
vụ rong biển của 2 khu vực này cũng chênh lệch. Nhìn chung, rong phía nam tồn
tại lâu hơn nên mùa vụ sinh học của rong biển kéo dài hơn.
Các loài rong biển phân bố chủ yếu ở vùng triều trở xuống. Sự phân bố
của rong biển phụ thuộc vào đặc tính cấu tạo, sinh lý, sinh thái của từng loài rong.
- Tầng cao triều: thành phần loài ít, chỉ những loài sống bám chắc vào vật
bám, có khả năng chịu khô hạn dài. Thường gặp là rong lục, rong mứt.
- Tầng triều giữa: rong biển phát triển mạnh, hình thành đai và những hợp
quần. Thường gặp rong lục, rong nâu như Chaetomorpha, Enteromorpha.
- Tầng triều thấp: rong có kích thước lớn, thích nghi ánh sáng mạnh,
thường gặp các loài rong nâu, rong đỏ và một số rong lục như Sargassum, Caulerpa, Padina.
V. MỘT SỐ LOÀI RONG BIỂN CÓ GIÁ TRỊ 1. Rong Câu Gracilaria 1.1. Phân bố
Gracilaria phân bố ở vùng khơi, nơi có độ mặn cao đến vùng cửa sông có
độ mặn biến động, có thể bắt gặp rong Câu ở vùng biển cũng như trong ao nước
tĩnh. Chúng phân bố từ vùng cao triều đến hạ triều và dưới triều.
Trên thế giới Gracilaria phân bố ở các vùng biển nhiệt đới, cận nhiệt đới và ôn đới.
1.2. Hình thái, cấu tạo
Hình thái: thân rong thẳng, có dạng trụ tròn hay dẹp. Bàn bám dạng đĩa.
Rong phân nhánh kiểu mọc chuyền, chạc hai, mọc chùm. Một số loài (G.
eucheumoides) có thân dẹp, mọc bò và tạo thành các bàn bám phụ từ mép các
nhánh. Một số loài khác thân có dạng lưỡi mác (ví dụ G. textorii). Cấu tạo:
Giải phẫu thân chính: các lớp của vỏ, kích thước, số lượng tế bào lõi, sự
thay đổi của tế bào từ vỏ đến lõi là những căn cứ phân loại đến loài.
Túi bào tử 4 phân bố dày trên bề mặt vỏ. Mỗi túi bào tử 4 gồm 4 bào tử
được xếp theo hình chữ thập.
Túi tinh tử: hình cầu hoặc hình oval, phân bố trên bề mặt thân. Vị trí và
dạng phòng tế bào của túi tinh tử cũng là căn cứ phân loại đến loài. Có 3 dạng:
Dạng 1: túi tinh tử phân bố trên bề mặt tản liên tục hoặc gián đoạn bởi các tế bào vỏ.
Dạng 2: túi tinh tử ở trong các phòng tế bào ở vị trí cạn.
Dạng 3: túi tinh tử ở trong các phòng tế bào sâu hơn, dạng trứng hoặc elip
kéo dài theo mặt cắt dọc. 49
1.3. Sinh sản và vòng đời
Sinh sản: gồm 3 hình thức, sinh sản dinh dưỡng, sinh sản vô tính, sinh sản hữu tính.
Vòng đời: trong vòng đời của Gracilaria, cây bào tử và cây giảo tử xuất
hiện luân phiên nhau. Cây bào tử 4 thành thục (2n) sinh sản (giảm phân) cho ra
các bào tử 4, chúng phát triển thành cây giao tử đực và cái. Cây giao tử đực
thành thục hình thành túi tinh tử, cây giao tử cái hình thành túi trứng. Sau khi thụ
tinh, cystocarp được hình thành trên cây giao tử cái, bào tử quả (2n) được phóng
ra và phát triển thành cây bào tử 4. Dạng cây dinh dưỡng của cây bào tử 4, cây
giao tử đực và cây giao tử cái không có khác biệt rõ ràng. 2. Rong thạch (Gelidium) 2.1. Phân bố
Rong bám trên đá, vỏ động vật thân mềm ở vùng trung triều đến hạ triều
và vùng nước sâu trên 10m. Chúng thích ứng với những nơi nước trong và dòng
chảy nhanh. Mùa vụ xuất hiện là xuân hè. Ở Việt Nam, rong thạnh phân bố từ
bắc vào nam. Trên thế giới, chúng phân bố ở vùng nước ấm, dọc bờ biển Thái
Bình Dương, Đại Tây Dương. Các loài phổ biến là G. amansii, G. pacificum, G.
divaricatum, G. crinale, G. japonicum…
2.2. Hình thái, cấu tạo
Hình thái: tản Gelidium dẹp, thẳng và rậm. Phân nhánh kiểu lông chim 4-
5 lần, có các nhánh nhỏ mọc cách hoặc mọc đối. Bàn bám dạng rễ giả. Kích
thước thay đổi đáng kể theo loài (từ 1-2cm đến 30-40cm). Rong thạch có màu đỏ
tía, đỏ nhạt hoặc nâu vàng (ở vùng biển nghèo dinh dưỡng).
Cấu tạo: Gồm hai cấu trúc cơ bản là lõi và vỏ. Các tế bào ngoài cùng ở
tầng vỏ sắp xếp thành hàng, mỗi tế bào có một sắc lạp, khoảng trống giữa các tế
bào được lấp đầy bởi các sợi nhỏ. Bên ngoài tầng vỏ được phủ đầy bởi một lớp
màng dày. Tầng lõi chủ yếu được tạo thành từ hàng chục nhóm tế bào trụ kéo dài
song song trục thân. Ở những phần mềm của tản, có một khoảng trống tương đối
lớn chứa đầy chất keo. Vì vậy Gelidium chứa hàm lượng keo rất cao. Ở những
tản thành thục, nhiều tế bào sợi chạy dọc xuyên qua chất keo làm cho mô của Gelidium dai và chặt hơn.
2.3. Sinh sản và vòng đời
Sinh sản: Rong thạch cũng có 3 hình thức thức sinh sản: sinh sản dinh
dưỡng, sinh sản vô tính, sinh sản hữu tính.
Sinh sản dinh dưỡng: gồm các hình thức sau:
- Sinh sản bằng thân bò: từ gốc của thân bò non hoặc tản trưởng thành nảy
những chồi bên. Chúng kéo dài theo chiều ngang tạo thành thân bò. Thân bò
phát triển xuống dưới và lên trên tạo thành rễ giả và chồi thẳng. Giống như cây
mây, thân bò kéo dài để tạo thành chồi thẳng liên tục.
- Sinh sản bằng cách mọc mầm: gồm có mọc mầm ở gốc của tản và mọc
rễ giả. Đó là các mầm nhỏ mọc từ gốc và rễ giả của cây non, từ đó phát triển
thành những cá thể rậm rạp.
- Sinh sản bằng đứt đoạn: sự đứt đoạn từ một cây mẹ cho ra những đoạn
rong. Chúng phát triển thành cá thể mới. 50
Sinh sản vô tính: trên đỉnh các nhánh nhỏ, có nhiều nhánh túi bào tử 4
dạng hình gậy. Tế bào mẹ túi bào tử bốn phát triển từ một tế bào vỏ trên đỉnh
nhánh nhỏ. Nó phân cắt tạo thành 4 bào tử bốn, sắp xếp theo dạng chữ thập hoặc
hình tứ diện thông qua quá trình giảm phân.
Sinh sản hữu tính: các tế bào vỏ kéo dài, phát triển thành tế bào mẹ túi
giao tử. Chúng phân cắt tạo thành những túi giao tử hình oval ở đỉnh nhánh. Sau
khi thành thục, túi giao tử sẽ sản xuất tinh tử. Còn các nhánh túi trứng ở cây giao
tử cái. Chúng phát triển từ những chồi nhỏ trên nhánh chính hoặc các nhánh bên.
Sau khi thụ tinh, tế bào túi trứng biệt hóa và phân cắt thành một hoặc nhiều túi
bào tử quả. Các tế bào vỏ ở bên ngoài túi bào tử quả giãn và nhô ra trên bề mặt
nhánh và tạo thành các cystocarp. Nhìn theo mặt cắt dọc, cystocarp có 2 khoang,
2 lỗ mà qua đó bào tử quả được phóng thích.
3. Rong sụn (Kappaphycus)
* Hệ thống phân loại: Ngành Rhodophyta Lớp Florideae Bộ Gigartinales Họ Solieriaceae Giống Kappaphycus 3.1. Phân bố
Vùng nhiệt đới châu Á, nhất là vùng tây Thái Bình Dương, đặc biệt ở
Philippines, Indonesia. Rong sụn phát trên rạn nơi có chất đáy cát - san hô, có
lưu chuyển nước ở mức trung bình, và nằm ở mức trung triều đến dưới triều.
Rong sụn Kappaphycus alvarezii hiện được trồng tại Việt Nam là loài
rong biển nhiệt đới, có nguồn gốc từ Philppines, được Phân viện khoa học Vật
liệu tại Nha Trang nhập nội và trồng từ tháng 2 năm 1993. Sự sinh trưởng và
phát triển của rong sụn chịu tác động trực tiếp của các yếu tố sinh thái môi trường chủ yếu sau:
- Độ mặn: Rong sụn là loài ưa mặn, chỉ sinh trưởng và phát triển ở các
vùng nước có độ mặn cao và tương đối ổn định, tốt nhất là từ 30 ‰ trở lên, thấp
hơn 20 ‰ và kéo dài nhiều ngày sẽ làm cho rong sụn ngừng phát triển và chết dần.
- Dòng chảy: Rong sụn phát triển ở các vùng nước có dòng chảy hay sự di
chuyển của dòng nước thông thoáng và thường xuyên. Nước bị tù hay di chuyển
kém làm cho sự phát triển của rong sụn chậm lại, đặc biệt nếu kết hợp với nhiệt
độ nước cao, chất huyền phù trong nước lớn, hàm lượng các muối dinh dưỡng
trong nước thấp sẽ dẫn đến sự tàn lụi của cây rong.
- Nhiệt độ: Rong sụn sinh trưởng tốt ở nhiệt độ từ 200C trở lên. Nhiệt độ
thích hợp cho rong sụn sinh trưởng và phát triển nằm trong khoảng 25 - 280C;
nhiệt độ cao hơn 300C và nhất là thấp hơn 200C sẽ ảnh hưởng xấu đến sinh
trưởng của cây rong. Nhiệt độ thấp hơn 15 - 180C thì rong ngừng phát triển.
3.2. Hình thái, cấu tạo
Có 3 dạng rong sụn hiện được nuôi trồng thương phẩm. Những loại rong
này khác nhau về hình thái. Chúng có thể dài, phân nhánh rời rạc với vài nhánh
phụ có đầu nhọn hoặc tù. Các nhánh phụ phân nhánh không theo quy tắc nào cả 51
và không hình thành dạng toả tròn như ở rong sần Eucheuma denticulatum. Cơ
thể có cấu tạo đa trụ.
3.3. Sinh sản và vòng đời
* Sinh sản: Chủ yếu là sinh sản hữu tính, vô tính.
* Vòng đời: Tương tự như rong câu và rong sần, cây bào tử và cây giao tử
của Kappaphycus xảy ra luân phiên trong vòng đời. Cây bào tử bốn thành thục
(2n) sinh sản (giảm phân) cho các bào tử bốn, chúng phát triển thành cây giao tử
đực và cái. Cây giao tử đực thành thục hình thành túi tinh tử, cây giao tử cái hình
thành túi trứng. Sau khi thụ tinh và cystocarp được hình thành trên cây giao tử
cái, bào tử quả (2n) được phóng ra và phát triển thành cây bào tử bốn.
4. Rong bẹ (Laminaria)
* Hệ thống phân loại: Ngành Phaeophyta Lớp Phaeosporeae Bộ Laminariales Họ Laminariaceae Giống Laminaria Loài Laminaria japonica 4.1. Phân bố
Loài L. japonica (Aresch) là loài có giá trị kinh tế nhất trong giống
Laminaria mặc dầu giống này có hơn 50 loài trên thế giới và khoảng 20 loài ở
vùng châu Á - Thái Bình Dương.
L. japonica phân bố ở vùng nước lạnh ôn đới. Ở khu vực châu Á - Thái
Bình Dương chúng phân bố về phía Nam đến vĩ độ 360 Bắc (Shangdong trở lên);
cho đến nay được nuôi trồng đến 250 Bắc (Fujian) vì ảnh hưởng lớn của nhiệt độ.
4.2. Hình thái, cấu tạo * Hình thái:
Cây rong dài 2 - 6 m, rộng 35 - 50 cm lúc trưởng thành. Bề mặt khum vào
lộ ra với ánh sáng. Trong tự nhiên, L.japonica sống đến 3 năm, trong nuôi trồng
người ta thu hoạch hàng năm: 8 tháng/vụ. * Cấu tạo:
- Phiến: Gồm 3 cấu trúc cơ bản.
+ Lớp vỏ ngoài: Là lớp mô ngoài cùng bao gồm các tế bào dậu và tế bào
nhỏ sắp xếp đều đặn, cạnh nhau. Tế bào dậu là những tế bào hình vuông theo
mặt cắt ngang, hình trụ dài theo mặt cắt dọc. Tế bào sắc tố tập trung gần bề mặt
hướng ra ngoài của tế bào vỏ.
+ Tầng bì: Nằm dưới lớp vỏ ngoài, biệt hoá thành ngoại bì và nội bì. Tế
bào tầng ngoại bì nhỏ hơn tế bào tầng nội bì. Cả hai tầng đều gồm những tế bào hình trụ kéo dài.
+ Tầng lõi: Gồm hai dạng tế bào, đó là tế bào lõi và tế bào sợi dạng kèn.
Cả hai dạng tế bào này đều nhỏ hơn nhiều so với tế bào nội bì. Tế bào tuyến keo
thường có dạng hình trụ. Các tế bào dạng kèn thì rất dài và có phần cuối dạng
kèn, chúng nối với nhau tạo thành mạng lưới các tế bào dẫn truyền. Cuối những
vách nơi mà những tế bào kèn nối với nhau có những đĩa dạng rây thủng nhiều 52
lỗ. Mạng lưới tế bào kèn cho phép các sản phẩm dự trữ chuyển từ nơi này sang
nơi khác. Chúng hoạt động như hệ thống mạch ở thực vật bậc cao.
- Các tuyến keo: Phân bố khắp tầng nội bì, cuống, bàn bám.
Cấu trúc của vách tế bào: Vách tế bào có hai lớp, lớp sợi và lớp gian bào
không định hình.Lớp sợi chủ yếu gồm cenllulose, chúng tạo thành khung cấu
trúc vững chắc của vách tế bào. Lớp gian bào tạo thành thể gốc (chất cơ bản) mà
các tế bào sợi được giữ chắc trong đó.
4.3. Sinh sản và vòng đời
* Sinh sản: Chủ yếu sinh sản hữu tính, vô tính.
* Vòng đời: Cây bào tử thành thục chứa túi bào tử nằm gần gốc của phiến.
Trong quá trình sinh sản vô tính đã xảy ra hiện tượng giảm phân ở các túi bào tử,
do đó các bào tử động có bộ nhiễm sắc thể n. Khi chín muồi, các bào tử động
được phóng thích ra khỏi cây mẹ. Chúng phát triển hình thành nên cây giao tử
đực và cây giao tử cái. Trên cây giao tử đực thành thục có những túi tinh tử, và
cây giao tử cái có những túi trứng. Trong túi tinh tử có chứa các tinh tử, trong túi
trứng có chứa trứng. Khi cây giao tử thành thục, tinh tử được phóng ra kết hợp
với trứng tạo thành hợp tử. Hợp tử qua quá trình phát triển hình thành nên cây bào tử. 53
CHƯƠNG 5. CỎ BIỂN
Cỏ biển là thực vật bậc cao có hoa, trái, lá và hệ thống mạch dẫn thật sự,
sống trong môi trường nước biển, có thể thụ phấn nhờ nước. Chúng chủ yếu
phân bố ở vùng nước nông gần bờ, chịu được tác động của sóng nhờ vào hệ
thống rễ ngầm. Cỏ biển thích nghi rộng với các điều kiện thủy lý, thủy hóa và
thủy sinh. Các thảm cỏ biển bao phủ một số vùng rộng lớn ở dải ven bờ với
nhiều chức năng lý – sinh học tạo nên một hệ sinh thái đặc thù.
Hiện nay, trên thế giới có khoảng 58 loài cỏ biển thuộc 12 chi, 4 họ và 2
bộ. Vùng Ấn Độ- Tây Thái Bình Dương là trung tâm về sự đa dạng của cỏ biển.
Vùng biển ven bờ Malaysia, Indonesia, New Guinea và bắc Australia có số
lượng loài cỏ biển được tìm thấy nhiều nhất.
Cỏ biển thuộc ngành Anthophyra, lớp Monocotyledoneae, bộ Helobiae.
Cỏ biển đã trở thành đối tượng nghiên cứu của các nhà khoa học trên thế giới
trong thập niên 80 của thế kỷ XX do vai trò của chúng đối với hệ sinh thái ven
biển của hầu hết các quốc gia (trừ các quốc gia ở vùng cực).
Cỏ biển được quan tâm nghiên cứu ở 2 khía cạnh quần xã và hệ sinh thái.
Thảm cỏ biển là nơi cư trú và cung cấp thức ăn cho nhiều loài sinh vật thủy sinh.
Môi trường biển luôn biến động bởi hoạt động của sóng gió và dòng chảy. Sự có
mặt của hệ cỏ biển làm giảm tác động cơ học của sóng, tạo môi trường thuận lợi
cho các sinh vật khác sinh sống. Lá của cỏ biển có vai trò “lọc nước”, bẫy và
lắng đọng trầm tích. Hệ thống rễ và thân ngầm chằng chịt của cỏ biển có tác
dụng cố định nền đáy, chống xói lở, góp phần làm ổn định đường bờ. Nhiều
công trình nghiên cứu cho thấy thảm cỏ biển vùng nhiệt đới là hệ sinh thái có
năng suất cao. Vì vậy, cùng với hệ sinh thái rừng ngập mặn và rạn san hô, cỏ
biển có vai trò quan trọng đối với vùng nước ven bờ, thực hiện các chức năng về
sinh học và cơ học. Các thảm cỏ biển tham gia vào chu trình chu chuyển vật chất
và chuỗi thức ăn ở vùng ven bờ. Nguồn vật chất hữu cơ được phân hủy từ lá cỏ
biển là thành phần quan trọng trong cơ sở thức ăn của hệ sinh thái biển.
Mục đích của việc nghiên cứu cỏ biển là phải xác định được nhiệm vụ của
hệ sinh thái và vai trò của chúng trong việc bảo vệ nguồn lợi thủy sản cũng như
trạng thái nguyên vẹn và ổn định của vùng biển ven bờ. Sự phát triển của cỏ biển
phản ánh trạng thái của vùng ven bờ và các thay đổi của chúng sẽ làm ảnh
hưởng đến môi trường và nguồn lợi sinh vật.
1. Đặc điểm hình thái
Các loài cỏ biển có hình dạng và kích thước rất khác nhau nhưng đều
mang nét độc đáo chung của các loài thực vật bậc cao trong nước: chúng thường
xuyên chịu tác động của sóng, gió, dòng chảy... vì vậy tất cả các loài cỏ biển đều
có thân ngầm, có hệ rễ chằng chịt trong nền đáy, giúp chúng có thể tồn tại lâu
năm trong khi phần thân đứng và lá có thể bị chết. Ví dụ phần thân ngầm của
loài Enhalus acroides ở Khánh Hòa có chiều dài 40-50cm và tồn tại trong vài chục năm. 2. Phân loại 54 Ngành Anthophyra Lớp Monocotyledoneae Bộ Helobiae
Hiện đã phát hiện được 15 loài cỏ biển ở Việt Nam: STT Tên loài Môi trường sống I Họ Hydrocharitaceae 1 Enhalus acroides
Vịnh kín, cửa sông, đầm, nơi có đáy bùn hoặc bùn cát 2 Halophila ovalis
Phân bố rộng, vùng triều, ven đảo, vịnh, ao, đầm, cửa sông, rừng ngập mặn 3 Halophila minor Vùng triều, vùng vịnh 4 Halophila beccarii Ao đầm nước lợ 5 Halophila sp Đầm có đáy bùn cát 6 Halophila decipiens
Vùng triều đáy cát hoặc bùn cát 7 Thalassia hemprichii
Vùng triều có đáy cát bùn hoặc có lẫn san hô chết II Họ Cymodoceaceae 8 Cymodocea serrulata
Vùng triều, vũng, vịnh ít sóng gió 9 Cymodocea rotundata
Vùng triều, vũng, vịnh, cửa sông 10 Halodule univervis
Vùng triều, vũng, vịnh, ao, đầm, rừng ngập mặn 11 Halodule pinifolia Vùng triều, đầm, phá 12 Ruppia maritima
Ao đầm rộng muối, rừng ngập mặn
13 Syringodium isoetifolium Vùng triều, ven đảo
14 Thalassodendron ciliatum
Vùng triều có đáy cát lẫn san hô chết III Họ Zosteraceae 15 Zostera japonica Vùng vịnh đáy cát bùn 3. Phân bố
Khác với rong biển, cỏ biển tồn tại và phát triển quanh năm. Chúng có thể
tồn tại nhiều năm với hệ thống thân ngầm và rễ chằng chịt tạo nên các cánh đồng rộng lớn.
* Phân bố theo các điều kiện sinh thái
Sự tồn tại và phát triển của các loài cỏ biển phụ thuộc chặt chẽ vào các
nhân tố môi trường mà quan trọng nhất là độ muối, nhiệt độ, độ đục, độ sâu và
cỡ hạt trầm tích. Ví dụ Halophila spinulosa và Thalassodendron coliatum thích
nghi vùng nước trong và sâu 2-17m. các quần thể địa phương Halophila bacearii
và Ruppia maritima chỉ được ghi nhận ở vùng nước lợ. Ngược lại Halophila
minor lại được thu thập trên nền đáy cát ở độ sâu tới 17m. Sự đa dạng loài cỏ
biển chịu ảnh hưởng của các nhân tố tại chỗ. Số loài nhiều nhất được ghi nhận ở
vùng có nền đáy bùn cát, được che chắn một phần tác động mạnh của sóng gió. 55
Ngược lại thành phần loài rất nghèo ở vùng đối sóng với nền đáy cứng hoặc
không ổn định và những nơi hoàn toàn bị che chắn với nền đáy bùn.
a. Cỏ biển ở vùng nước triều và ven các đảo
Ở vùng triều, nước trong, sóng đánh mạnh, độ mặn cao nền đáy là san hô
chết lẫn với cát, đáy dốc thẳng và tương đối bằng phẳng có các loài Thalassia
hemprichii, Cymodocea spp, thường chiếm ưu thế ở độ sâu tối ưu từ 0-2m. Lát
cắt ngang qua các vùng triều thấp là các loài Cymodocea rotundata, Cymodocea
serrulata, Halodule univervis trên đáy cát bùn. Tiếp đến là Thalassia hemprichii
và Halophila sp. Ở các nơi khuất sóng gió có loài Enhalus acroides nhưng mật
độ thưa và kích thước cá thể nhỏ. Ngoài ra, ven các đảo (Côn Sơn, Phú Quốc...)
trên các bãi bằng phẳng và tương đối ít sóng, từ vùng triều thấp trở xuống, trong
quần xã cỏ biển còn có thêm loài Syringodium isoetifolium.
b. Cỏ biển ở vùng đầm, vịnh
Trong đầm, vịnh kính và yên sóng, đáy bùn hoặc bùn cát như ở đầm Thủy
Triều, vịnh Cam Ranh (Khánh Hòa), đầm Cù Mông (Phú Yên), phá Tam Giang
(Thừa Thiên Huế)... cỏ biển phân bố thành vùng rộng lớn (có thể lên đến 800 ha
như ở vịnh Cam Ranh). Sự đa dạng loài cỏ biển ở đây là cao nhất, hầu hết các
loài cỏ biển đều hiện diện trong môi trường này. Từ Đà Nẵng trở ra còn gặp
thêm loài Zostera japonica. Vào mùa mưa lũ, độ mặn trong đầm giảm xuống đột
ngột, phần lá dễ bị thối rữa nhưng phần thân ngầm vẫn tồn tại và sẽ phát triển trở
lại khi mùa mưa chấm dứt. Loài ưu thế và quan trọng nhất là Enhalus acroides.
Chúng mọc thành các đồng cỏ dày, trung bình cao khoảng 1m. Đây là nơi trú ẩn,
nuôi dưỡng nhiều loài hải sản có giá trị, có vài trò quan trọng đối với việc duy trì
và bảo vệ nguồn lợi sinh vật và môi trường.
c. Cỏ biển trong các ao, rừng ngập mặn, cửa sông
Ở trong các ao, đìa nuôi trồng thủy sản, rừng ngập mặn hoặc ruộng muối
bỏ hoang, có các loài cỏ biển kích thước nhỏ như Halophila sp, Ruppia
maritima, Halodule univervis. Loài Halophila ovalis thích nghi với mật độ cao
(trên 10.000 cây/m2) tạo thành thảm cỏ dày phủ kín nền đáy, cao vài cm. Ở vùng
cửa sông còn gặp các loài Cymodocea rotundata và Halodule univervis. Đặc biệt
các loài Halophila beccarii và Ruppia maritima chỉ xuất hiện ở fđây mà không
gặp ở vùng triều ven biển.
* Phân bố theo độ sâu
Các loài cỏ biển phân bố từ vùng triều thấp trở xuống, đặc biệt là từ mức
triều thấp nhất đến độ sâu 3-4m. Halophila sp mọc ở độ sâu nhỏ còn Halophila
ovalis, Cymodocea serrulata, Halodule univervis thích nghi rộng về độ sâu. Ở
vùng triều và ven các đảo, nước trong chúng có thể mọc ở độ sâu 10-20m.
4. Các quần xã cỏ biển
Ở những nơi có điều kiện tự nhiên thích hợp, các loài cỏ biển thường mọc
tập trung thành các quần xã hỗn hợp nhiều loài. Ngược lại, một số nơi có điều
kiện sinh thái đặc biệt, chỉ có một loài thích nghi, chúng sẽ phát triển ưu thế tạo
thành các quần xã thuần loại, xâm chiếm, che phủ toàn bộ nền đáy. Một vài loài 56
khác có thể phát triển trên các vùng rộng lớn hàng trăm ha hoặc tạo thành những vành đai hẹp.
1. Quần xã hỗn hợp Thalassia hemprichii và Halophila ovalis
Ở vùng triều ven biển, loài cỏ Vích Thalassia hemprichii và loài cỏ Xoan
Halophila ovalis mọc xen kẽ, rải rác trên đáy cát lẫn san hô chết. Loài cỏ Vích
thường chiếm ưu thế, loài cỏ Xoan mọc thưa hơn. Quần xã này xuất hiện phổ
biến ở vùng dưới triều ven biển Khánh Hòa, Ninh Thuận.
2. Quần xã hỗn hợp Enhalus acroides, Thalassia hemprichii và Cymodocea serrulata
Các loài này mọc chung với nhau ở vùng triều nơi khuất sóng, tạo thành
thảm dày che kín nền đáy. Loài Enhalus acroides mọc thành từng đám, xen giữa
những đám đó là Thalassia hemprichii và Cymodocea serrulata. Kiểu quần xã
này thường gặp ở Ninh Thuận, Khánh Hòa.
3. Quần xã hỗn hợp Cymodocea rotundata, Halophila ovalis và Halodule univervis
Các loài này mọc xen kẽ nhau trên đáy cát bùn ở các vịnh, cửa sông nhỏ.
4. Quần xã Enhalus acroides
Môi trường sống thích hợp của loài này là các vũng, vịnh có đáy bùn hoặc
bùn cát. Chúng cũng được bắt gặp ở vùng triều ven biển khuất sóng có đáy cát
lẫn mảnh vụn san hô nhưng mật độ và sinh vật lượng kém. Do kích thước cá thể
lớn và có diện tích phân bố rộng nên các đồng cỏ này có tính đa dạng sinh học
cao, tạo nên vùng khai thác quan trọng của nghề cá ven bờ.
5. Quần xã Thalassia hemprichii
Loài Thalassia hemprichii chiếm ưu thế ở những bãi triều tương đối bằng
phẳng ven biển, ven đảo có đáy cát lẫn san hô chết. Chúng tạo thành những đám
cỏ lớn, có mật độ cao như ở đảo Phú Quý (Bình Thuận), Khánh Hòa, Côn Sơn...
Đây là quần thể đặc trưng cho vùng triều ven biển, là nơi cư trú của nhiều loài
sinh vật có giá trị ở vùng triều.
6. Quần xã Halophila ovalis
Loài Halophila ovalis thích nghi rộng với các điều kiện môi trường.
Chúng có tốc độ sinh trưởng nhanh và thường xâm chiếm nền đáy ao nuôi tôm,
đầm, vịnh hoặc các kênh mương dẫn nước mặn. Mật độ thảm cỏ này thường trên 20.000 cây/m2.
7. Quần xã Halophila beccarii
Halophila beccarii thường xuất hiện trong các kênh mương, ao nước lợ,
rừng ngập mặn, tạo thành thảm dày với mật độ trên 30.000 cây/m2.
8. Quần xã Halodule univervis
Halodule univervis thích nghi nhất ở các đầm vịnh. Lá có thể dài đến
20cm, mọc phủ kín nền đáy giống như cỏ trên lục địa. Chúng phân bố từ vùng
triều thấp đến độ sâu hơn 10m. 57
9. Quần xã Ruppia maritima
Ruppia maritima thường gặp trong các ruộng muối, ao nuôi tôm, rừng ngập mặn.
5. Vai trò của cỏ biển
Cỏ biển được quan tâm ở 2 khía cạnh: quần xã và hệ sinh thái. Về quần
xã, các đồng cỏ biển là nơi mà các quần thể động vật, thực vật có mối quan hệ
tương hỗ sinh sống. Về hệ sinh thái, các diễn biến của mối quan hệ này được
kiểm soát bằng tác động qua lại của các yếu tố sinh học của môi trường.
- Vai trò sinh thái của thảm cỏ biển được quyết định bởi tốc độ tạo thành
hữu cơ nhanh chóng của chúng. Tính theo đơn vị diện tích, giá trị này cao hơn
năng suất của thực vật phù du ngoài khơi Pêru – vùng có năng suất sinh học cao
nhất thế giới. Nhờ sự cố định năng lượng mặt trời có hiệu quả và sản lượng sinh
khối cao, cỏ biển có khả năng tăng cường và duy trì độ phì nhiêu của thủy vực.
Điều này còn được bổ sung bởi quá trình trao đổi vật chất hữu cơ hiệu quả diễn
ra trên lá và nền đáy. Quá trình này được thực hiện bởi cấu trúc dinh dưỡng đa
dạng cao và được tăng cường bởi các chất nội tạicungx như ngược lại.
- Làm giảm năng lượng và chuyển động của nước biển
Thảm cỏ biển dày với hệ rễ neo chặt vào nền đáy có tác dụng làm giảm
năng lượng của sóng, dòng chảy. Nhờ vậy chúng có khả năng chống xói lở, bảo
vệ đường bờ, tạo nên vùng đệm chắn sóng. Các loài cỏ biển có kích thước lớn có
tác dụng làm giảm năng lượng và sự chuyển động của nước biển. Điều này có
hai tác dụng: tạo nơi trú ngụ, ẩn nấp cho sinh vật khác, còn đối với môi trường,
chúng làm gia tăng trầm tích, giảm sự xáo trộn và vẩn đục của các chất hữu cơ
cũng như vô cơ trong nước.
- Bẫy, giữ và cố định trầm tích
Tác động của cỏ biển trong việc bẫy, giữ trầm tích phụ thuộc vào loài cỏ
và mật độ của chúng. Hệ thống rễ và thân ngầm có ý nghĩa lớn nhất về phương
diện này nhưng lá có vai trò quyết định do chúng tác động vào sự chuyển động
của nước. Ví dụ Enhalus và Thalassia tác động mạnh mẽ hơn Halodule hoặc
Halophila bởi hệ thống rễ và thân ngầm rất phát triển, lá to và dài. Việc bẫy và
cố định trầm tích được biểu thị bởi sự tạo nên các gò liên tiếp dưới biển. Ở
những vùng chịu nhiều bão tố, cỏ biển có vai trò lưu giữ trầm tích nhờ hệ thống
thân, rễ ngầm. Cấu trúc thân, rễ và đặc tính tăng trưởng phản ánh đặc tính của
chế độ trầm tích thủy vực. Mặt khác thảm cỏ biển là bộ máy có hiệu quả cao đối
với việc hấp thụ chất dinh dưỡng, chất thải từ đất liền và có vai trò như những
bẫy trầm tích làm giảm độ đục của nước.
- Cung cấp nơi sinh sống và là cầu nối trong con đường di cư của nhiều sinh vật
Về phương diện này, cỏ biển là nơi dự trữ đa dạng gen và có vai trò như
rừng ngập mặn. Chúng có thể đảm nhiệm một phần vai trò của rừng ngập mặn ở
những nơi rừng bị tàn phá. Do hình thể đa dạng của lá, thân đứng và các quần xã 58
rong biển sống chung, các đồng cỏ biển đã cung cấp nơi cư trú thích hợp và
nguồn dinh dưỡng đầy đủ cho các loài sinh vật biển.
Các thảm cỏ biển thường phát triển ở vùng trung gian của rừng ngập mặn
và rạn san hô hoặc là vùng đệm của hai hệ sinh thái khác. Vì vậy chúng trở thành
điểm dừng chân của nhiều loài cá, động vật không xương sống, thú và bò sát. Ví
dụ thảm cỏ tóc tiên phơi bãi khi triều thấp ở Bắc Đại Tây Dương là một phần
của con đường di cư của chim nước. Bằng việc cung cấp nơi ẩn náu thông qua
tán cây và hình thái, kích thước khác nhau của bóng khí cũng như nguồn dinh
dưỡng giàu có, thảm cỏ biển trở thành bãi ương giống chất lượng cao của nhiều
thủy sinh vật. Nguồn giống sau khi được nuôi dưỡng ở đây sẽ được phát tán đến
các hệ xung quanh của đại dương.
Qua hoạt động quang hợp, cỏ biển sử dụng năng lượng ánh sáng mặt trời
để tổng hợp nguồn chất hữu cơ cung cấp cho biển. Sức sản xuất của cỏ biển rất
cao (ví dụ loài Enhalus acroides có sức sản xuất sơ cấp là 0,95-2,31
gC/m2/ngày). Bên cạnh đó, cỏ biển còn là nguồn cung cấp oxy rất hiệu quả cho môi trường.
- Cung cấp cho loài người những sản phẩm trực tiếp như vật liệu di
truyền, thực phẩm; vật liệu thô cho công nghiệp và năng lượng
Ngoài ra, thảm cỏ biển được coi là môi trường thuận lợi cho nuôi trồng
trên biển, du lịch biển… đem lại lợi nhuận cao. 59
TÀI LIỆU THAM KHẢO
1. Nguyễn Hữu Đại, 1999, Thực vật thuỷ sinh, Nhà Xuất Bản Nông
Nghiệp Tp.HCM, Viện Hải Dương Học Nha Trang. 2. Lê Thanh Hùng, 2
000, Bài giảng dinh dưỡng và thức ăn Thủy sản
3. Đặng Đình Kim & Đặng Hoàng Phước Hiền, 1999, Công nghệ sinh học vi tả , o Trung tâm KHTN & CNQG. 60