Ôn tập sinh học phân tử | Đại học Y Dược Thành phố Hồ Chí Minh
Nghiên cứu cấu trúc và chức năng gen. Sản xuất protein và sản phẩm của gen bằng phương pháp tái tổ hợp. Tạo chủng VSV chuyển gen (GMO). Hoàn thành bản đồ gen người, điều trị một số bệnh bằng pp “trị liệu gen”. Kết hợp SHPT và Tin Học (In silico- nghiên cứu sinh học trên máy điện toán). Tài liệu giúp bạn tham khảo, ôn tập và đạt kết quả cao. Mời đọc đón xem!
Môn: Phương pháp và kỹ thuật cơ bản trong sinh họcphân tử 11 tài liệu
Trường: Đại học Y Dược Thành phố Hồ Chí Minh 379 tài liệu
Tác giả:
















Preview text:
lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
ÔN TẬP SINH HỌC PHÂN TỬ
Biên soạn: Nguyễn Đắc Nhân – DCQ2016
CHƯƠNG I: LỊCH SỬ PHÁT TRIỂN CỦA SINH HỌC PHÂN TỬ I. Tiên đề.
- 1869: Fried Miescher phát hiện acid nucleic.
- 1865: Mendel khám phá quy luật di truyền.
- 1982: Griffith ghi nhận chuyển đổi kiểu hình từ Rough (không gây bệnh)
sang Smooth (gây bệnh) ở Streptococcus pneumonias.
- 1944: Oswaid Avery và cộng sự công bố thí nghiện đầu tiên chứng minh
ADN là vật liệu di truyền.
- 1951: Chứng minh ADN có A=T, G=C.
- 1952: Hershey và Chase chứng minh ADN mang thông ti duy truyền bằng phương pháp phóng xạ.
- 1953: Watson-Dick xác định mô hình xoắn kép của ADN.
- 1961: Tìm ra bộ ba mã hóa di truyền đầu tiên UUU → Phenylalanin. Giữa
thập niên 60, toàn bộ 64 bộ ba mã hóa được xác định.
- 1967: Chiết xuất enzyme ligaza.
- 1962: Phát hiện enzyme cắt giới hạn trong TB VK.
- 1970: Chiết enzyme cắt giới hạn P.E. Restriction enzyme. → Tạo ra ADN
plasmid có khả năng tái tổ hợp, sao chép độc lập với NST.
- 1985: Saiki và Mullis sử dụng PCR để khuếch đại đoạn gen. Xác định trình
tự nucleotide → gây ĐB điểm ảnh hưởng → tạo các biến đổi tùy ý trên gen.
II. Ứng dụng SHPT.
- Nghiên cứu cấu trúc và chức năng gen.
- Sản xuất protein và sản phẩm của gen bằng phương pháp tái tổ hợp.
- Tạo chủng VSV chuyển gen (GMO).
- Hoàn thành bản đồ gen người → điều trị một số bệnh bằng pp “trị liệu gen”.
Kết hợp SHPT và Tin Học (In silico- nghiên cứu sinh học trên máy điện toán). III.
Học thuyết trung tâm.
“Thông tin di truyền đã chuyển sang Protein thì không thể lấy lại được.”
- Thông tin: Trình tự chính xác của các nu, axit nu → quy định trình tự aa →
TTDT không thể truyền ngược từ Protein đến ADN. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
IV. Các kỹ thuật dùng trong SHPT.
1. Tạo dòng biểu hiện.
- Dùng để nghiên cứu chức năng của Protein bằng cách tạo dòng ADN vào một
plasmid và đưa vào tế bào để biểu hiện.
- Nhờ cách thay đổi cơ cấu biểu hiện trên plasmid hoặc protein tinh chế, ta hiểu
được cấu trúc và chức năng của gen, cơ chế điều hòa của gen, quan hệ của nó
với hoạt động của các gen khác hay protein khác. 2. Kỹ thuật PCR.
- Kỹ thuật khuếch đại gen đặc hiệu để thu được nhiều bản sau.
- Cho phép dễ dàng đưa các thay đổi, đột biến vào ADN để nghiên cứu.
3. Kỹ thuật điện di gel.
- Đây là công cụ phân tích chính trong SHPT.
- Các phân tử ADN, ARN, Protein dưới tác động của điện trường xuyên qua một
hệ gel nào đó → tách nhau tùy theo kích thước hay điện tích.
4. Kỹ thuật Southern blot và Northern blot.
a. Kỹ thuật Southern blot.
- Kỹ thuật vết lai southern blot dùng để phát hiện sự hiện diện của một trình tự ADN đặc biệt.
- Phương pháp: Các ADN được phân tích trên gel và chuyển sang một màn rắn
→ phát hiện vết ADN bằng phép lai với đầu dò đặt hiệu → Xác định trình tự ADN đặc biệt.
b. Kỹ thuật Northern blot.
- Kỹ thuật Northern blot tương tự kỹ thuật southern blot.
- Nghiên cứu sự biểu hiện ARN khi các ADN được phân tích trên gel và chuyển
sang một màng rắn → phát hiện ARN bằng phép lai với đầu dò ARN.
5. Kỹ thuật Western và hóa miễn dịch.
- Protein được phân tích trên gel và chuyển sang các màng rắn lai vết miễn
dịch) → phát hiện protein trong tế bào bằng phương pháp lai và liên hệ với
sự biểu hiện của gen tương ứng. 6. Kỹ thuật Array
- Cho phép đặt rất nhiều mẫu dò acid nucleic vào các chấm nhỏ trên chip. Các
mẫu dò được thiết kế có trình từ bổ sung với acid nucleic (ADN hay ARN) đích.
- Array được dùng để nghiên cứu sự biểu hiện gen và có thê thiết kế sử dụng
kháng thể để nghiên cứu sự biến động của protein (protein array).
CHƯƠNG II: SAO CHÉP ADN I.
Cấu trúc phân tử ADN.
1. Cấu trúc bậc 1. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
Nu và trình tự sắp xếp các nu trên gen. Thành phần nucleotit bao gồm Bazo nito,
Axit photphoric và đường Deoxyribose (C5H10O4).
- Bazo nito: Loại lớn (purin) gồm Adenin và Guamin và loại nhỏ (pirimidin) gồm Timim và Cytosin.
- Nucleosic: Bazơ nitơ + C5H10O4. - Liên kết bazo.
2. Cấu trúc bậc 2.
Cấu dạng không gian 3 chiều, gồm 3 dạng cấu trúc bặc 2
- Dạng B: là dạng chủ ye6u1m tồn tại trong môi trường 92% H2O, cấu trúc
xoắn bàn phải (3’ – 5’ theo chiều ngón tau cái phải).
- Dạng A: Tồn tại trong môi trường 72% H2O, cấu trúc bàn tay phải.
- Dạng Z: Điều hòa hoạt động gen, cấu trúc bàn tay trái.
3. Cấu trúc bậc 3. Sắp xếp ADN
trong NST II. Sao chép ADN. 1.
Thí nghiệm Melselson và Stahl.
Thực hiện trên Vi khuẩn: Nuôi cấy vi khuẩn E.coli 15N trong môi trường 14N.
Khi chuyển sang môi trường 14N làm cho trong phân tử ADN gồm 1 sợi mới và 1 sợi cũ. 2. Sao chép ADN.
-Chỉ tạo đầu 3’ có 3’-OH tự do gắn với phosphin tự do tạo liên kết
phosphodieste nên sao chép luôn từ đầu 3’ theo chiều 3’ → 5’.
Phương trình: d(NMP)n+ d(NTP) → d(NMP)n+1 +
PPi d(NMP) : Deoxyribonucleosic monophosphat.
d(NTP): : Deoxyribonucleosic triphosphat. PPi: Pyrophosphat
* Thành phần tham gia vào quá trình sao chép ADN. - ADN khuông mẫu.
- dNTP: dA, dT, dG, dC. - ADN-polymerase - Mg2+. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
* Các kiểu sao chép ADN.
- Sao chép Theta → Vi khuẩn.
- Sao chép lăn vòng → Virut, yếu tố giới tính F của E.Coli.
- Sao chép thẳng → TB nhân thật.
3. Các loại enzyme tham gia vào quá trình sao chép ADN. a. Enzym Helicase.
- Enzym cắt đứt 2 mạch của ADN. - Đặc điểm cấu trúc:
+ Chuỗi axitamin có trình tự bảo thủ: giống nhau ở tất cả mọi loài. + Gắn ATP. + Thủy giải ATP. + Gắn axit amin.
- SSB protein: làm cho 2 sợi ADn không xoắn lại với nhau. b. Enzym ADN- polymerase.
- Vai trò polymer hóa và sửa sai ( exonuclease). NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ -
Đây là một loại holo enzyme gồm Apoenzym + Co-factor Mg2+ → Holoenzym
ADN polymerase ( hoạt tính) có khả năng sửa chữa hư hỏng trên sợ ADN mới
tổng hợp khi phát hiện sai sót. ADN-polymerase Polymer Exonuclease Chức năng hóa 5’ – 3’ 5’ – 3’ 3’ – 5’ I II + + + E.coli + - + III + + + Virut α β + - + α + + - Hoạt tính primerase β γ + + - Sửa sai, tái tổ hợp Tế bào nhân ADN thật + + + Sao chéo, sữa chữa ADN trong ty thể δ + + + Sao chép ADN ε + + + Chưa rõ c. Enzym ligase. - Enzym nối. d. Enzym Topoisomerase.
- Vai trò: điều hòa tình trạng siêu xoắn của ADN, gồm 2 loại +
Topoisomerase 1: cắt 1 sợi ADN, không cần ATP.
+ Topoisomerase 2: cắt 2 sợi ADN, cần ATP. e. Enzym primarase.
- Prumarase + polypeptide → primosome.
- ADN helicase kích hoạt primarase tổng hợp đoạn mồi ARN có chiều dài
11±1 nu → cung cấp 3’OH giúp ADN-polymerase gắn ADN. Trên sợi liên
tục có 1 mồi ARN tận cùng 5’ và trên sợi không liên tục có nhiều mồi
ARN, mỗi đoạn okazaki có 1 mồi.
4. Quá trình sao chép ADN ở TB nhân nguyên thủy – E.coli
a. Giai đoạn khởi đầu.
- Trình tự OriC ( dài khoảng 245 bp) cùng với protein khởi đầu tiến hành tháo xoắn đoạn ngắn ADN. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ -
- Enzim helicase cắt đứt hai mạch của chuỗi xoắn kép. SSB protein giữ cho hai
mạch không bị dính vào nhau. Isotopomerase tác động vào một mạch hoặc hai
mạch giúp hai mạch đơn ADN tách nhau ra.
b. Giai đoạn tổng hợp mạch mới.
- ADN-polymerase tiến hành tổng hợp đoạn mới theo chiều 5’ – 3’.
Trong quá trình tổng hợp mạch mới cần có enzim tổng hợp đoạn mồi
primerase ( kết hợp protein tạo primesome) dưới sự hoạt hóa của enzyme helicase. c. Giai đoạn kết thúc.
- Quá trình sao chép kết thúc khi: Hai chạc chữ Y gặp nhau, gặp protein kết thúc (
Tus) hoặc gặp trình tự kết thúc. Khi đó ADN-polymerase sẽ ngừng tổng hợp,
enzyme ligase tiến hành nối các đoạn okazaki và hai phân tử protein mới được giải phóng.
5. Quá trình sao chép ADN ở TB nhân thật.
- Quá trình sao chép ADN ở TB nhân thật diễn ra trên nhiều điểm sao chép (
nhiều replicon) → Sao chép nhanh hơn TB nhân nguyên thủy.
- ADN kết hợp với protein histon tạo nucleosome.
- Mỗi đoạn axit amin dài 40-300 nu. - Vận tốc trượt của ADN-polymerase 50nu/s.
6. Quá trình sao chép ADN Virut.
a. Cấu trúc TTDT của virut. - ARN sợi đơn - ADN + Sợi đơn.
+ Sợi đôi: Thẳng hoặc vòng.
b. Sao chép ADN ở phago T7.
- Hệ gen dsADn có 39937 bp.
- Vị trí khởi đầu sao chép cách vị trí cuối cùng trải 59000 bp.
- Đầu tận cùng phải và trái chứ trình tự thừ 160bp.
c. Sao chép ADN ở phago lamda
- Cơ chế sao chép tạo nhiều bản sao bằng hình thức lăn vòng.
- Ở mỗi tận cùng của bộ gen là một đoạn đơn dài 12 nucleotid và được gọi là vị
trí cos, hai đoạn dơn ở vị trí cos có thể gắn bổ sung với nhau để biến bộ gen thành dạng vòng dôi. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ -
- Nhờ trình tự cos làm cho NST có thể vòng hóa, tránh bị phân giải bởi enzyme
exonuclease của vi khuẩn và có thể được sao chép theo chu trình tiêu giải tiềm ẩn.
d. Sao chép ADN ở phago M13.
- Cơ chế sao chép lăn vòng tạo 1 bản sao. - Protein SSB bao bọc AD.
- Không tổng hợp được ADN polumerase riêng biệt nên quá trình sao chép
lệ thuộc tế bào vật chủ. Một phần của bộ gen có cấu trúc nút kẹp tóc sợi
đôi. Vùng này đóng vai trò promoter cho ARN-polymerase của tế bào vật
chủ bám vào và phiên mã một đoạn ARN ngắn. ADN polymerase-III stham
gia và tổng hợp sợi đôi ADN, gọi là thể sao chép I (RFI – replicative for I).
7. Sữa sai ADN không sao chép
Các enzyme trượt dò tìm và sửa các lỗi sai: Bazo biến đổi hóa học, liên kết cộng
hóa trị sai, bắt cặp sai. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
CHƯƠNG III: CÁC LOẠI ARN I. Khái niệm. 1. Cấu trúc ARN:
- Mạch đơn polynucleotide. - Đường Ribose.
- Bazo nito: A,G,C,U. → Khác so với ADN:
+ Đường ribose có thêm nhóm – OH nên kém bền hơn ADN.
+ Cấu trúc mạch đơn. + Bazo nito: T thay bằng U. II. Các loại ARN. - Gồm 03 loại ARN chính:
+ mARN: Chứa trình tự nucleotide từ ADN để mã hóa protein.
+ tARN: vận chuyển acid amin đến Ribosome.
+ rARN: là thành phần cấu tạo của Ribosome. - Các ARN khác:
+ pre – rARN( tiền tARN): Nhân → pre – rARN → cắt nối: rARN hoàn chỉnh.
+ pre-tARN ( tiền tARN): Nhân → pre-tARN → cắt nối: tARN hoàn chỉnh.
+ hnARN (heterogenous nuclear ARN): ARN nhân không đồng nhất → cắt nối: mARN.
+ snARN ( small nuclear ARN): ARN nhân nhỏ.
+ scARB ( small cytoplasmic ARN): ARN tế bào chất nhỏ. - Phân loại:
+ ARN mã hóa: hnARN → mARN – ARN mã hóa.
+ ARN không mã hóa: các ARN còn lại.
- Trong Tế bào, ARN ở trạng thái: + Tự do.
+ Liên kết Protein => nucleoprotein.
- Sản phẩm đầu tiên của phiên mã ở TB nhân thật là các hnARN: có các đoạn
Inxon, Exon xen kẽ. Một số TB Prokaryote cũng có gen gián đoạn. 1. rARN.
- Ribosome = các phân tử rARN + 50 protein. -
%mrARN = 50% mRb - Ba loại rARN ( phân theo độ lắng).
+ rARN ở TB Prokaryote: 70S = 50S (31 pro) + 30S (21 pro)
+ rARN ở TB Eukaryote: 80S = 60S ( 49 pro) + 40S ( 33 pro)
+ rARN ở Ty thể ĐV có vú : 50S
a. Cấu tạo Ribosome. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
- Gồm tiểu đơn vị lớn và tiểu đơn vị nhỏ (subunit).
- Gồm 1 hay nhiều rARN cùng nhiều phân tử protein.
- Các tiểu đơn vi Ribosome:
+ Được tổng hợp trong nhân.
+ Di chuyển ra tế bào chất qua lỗ nhân.
+ Hai tiểu đơn vị liên kết với nhau tại Tế bào chất. - Vai trò của Ribosome:
+ Tiểu đơn vị lớn: Chịu trách nhiệm hình thành liên kết peptide trong dịch mã.
+ Tiểu đơn vị nhỏ: xác định acid amin cần tổng hợp, kiểm soát quá trình tương
tác giữa tARN và anticodon trong dịch mã.
b. Cấu trúc rARN.
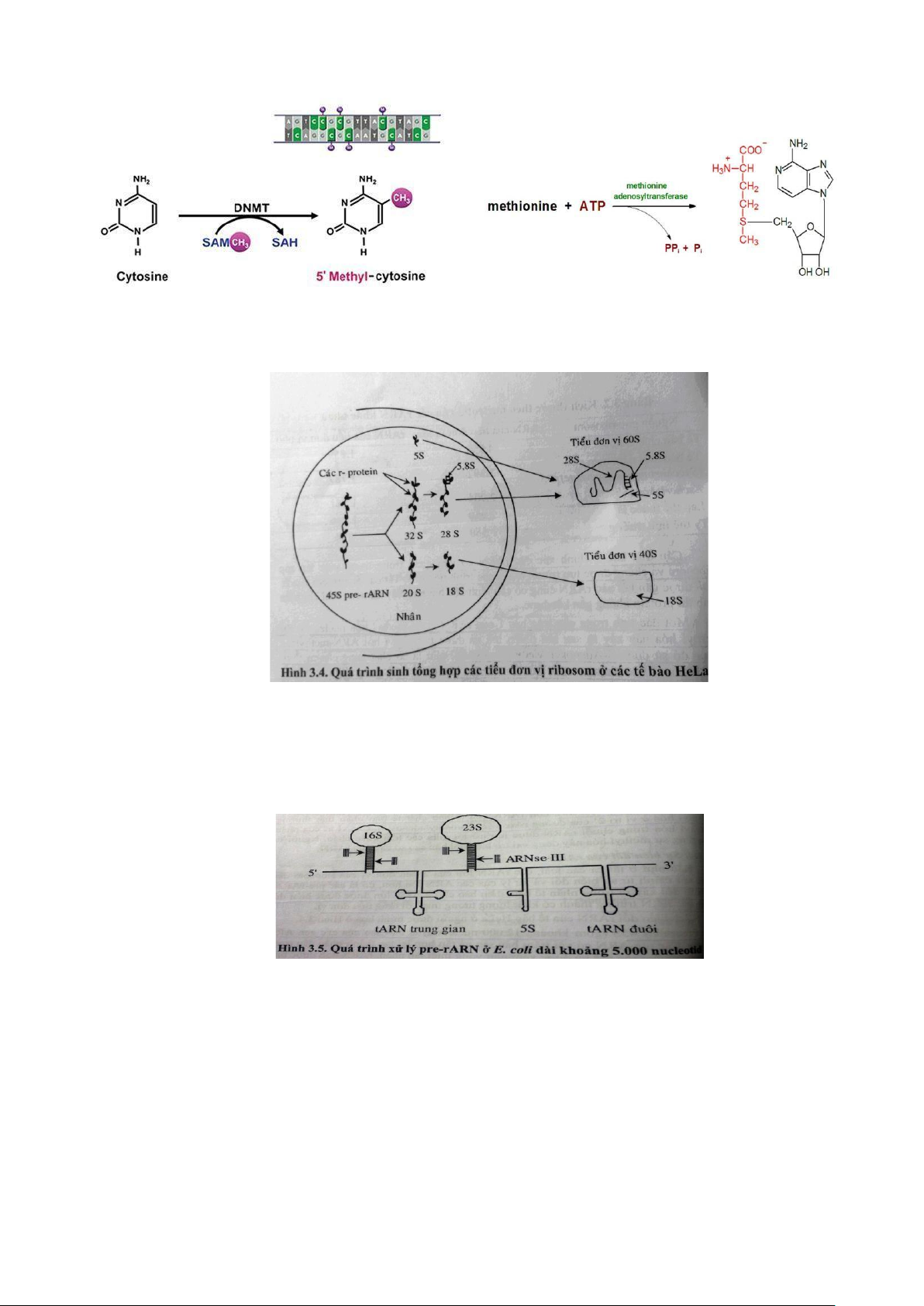
- Cấu trúc bậc I: methyl hóa nucleotide, biến đổi bàn sao sơ cấp.
+ Thực hiện bởi ARN – methylase.
+ Cơ chất cung cấp methyl: S- Adenosyl Methionin (SAM).
+ Methyl hóa: nhân sơ ở điểm khởi đầu (5-methyl citozin), nhận thật ở vị trí 2’- O-methyladenolsyl. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
- Cấu trúc bậc II: Lắp ráp ribosome.
+ TB Eukaryote ( TB nhân thật):
+ TB Prokaryote (TB nhân sơ): chuỗi pre-ARN có chứa hai hay nhiều hơn
tARN. Chuỗi 5S rARN là thành phần của đơn vị phiên mã của cả rARN 16S và rARN 23S.
• RNase III: cắt 16S rARN và 23S rARN.
• RNase E và M5: cắt 5S rARN. • Rnase P và F cắt ARN. 2. tARN. -
Vận chuyển acid amin đến mARN dựa trên anticodon và codon. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
a. Đặc điểm của tARN.
Kích thước nhỏ: 73-93 nu.
- Cấu trúc chiếc lá chẻ 3.
- Đầu tần cung 2luo6n kết thúc bằng trình
tự CCA – gắn kết acid amin. + E.coli: CCA
được phiên mã bổ sung có trong ADN nhiễm sắc thể,
+ TB Nhân thật: CCA được thêm vào sau khi phiên mã.
b. Cấu trúc của tARN. - Đầu tận cùng 5’P.
- Nhánh gắn các acid amin gồm 7 cặp base.
+ Các nucleotide tận cùng có 3’-OH tự do để gắn acid amin
+ Các cặp base có thể không theo trình tự bổ sung
- Đuôi CCA: trình tự CCA tại 3’ giúp
enzyme nhận biết được tARN khi dịch mã.
- Vòng D: gồm 4 cặp base, chứa dihydrouriridine.
- Vòng đối mã: gồm 5 cặp
base. Mỗi tARN mang một bộ ba đối mã
đặc hiểu của 1 hay nhiều codon mã hóa cho 1 acid amin. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
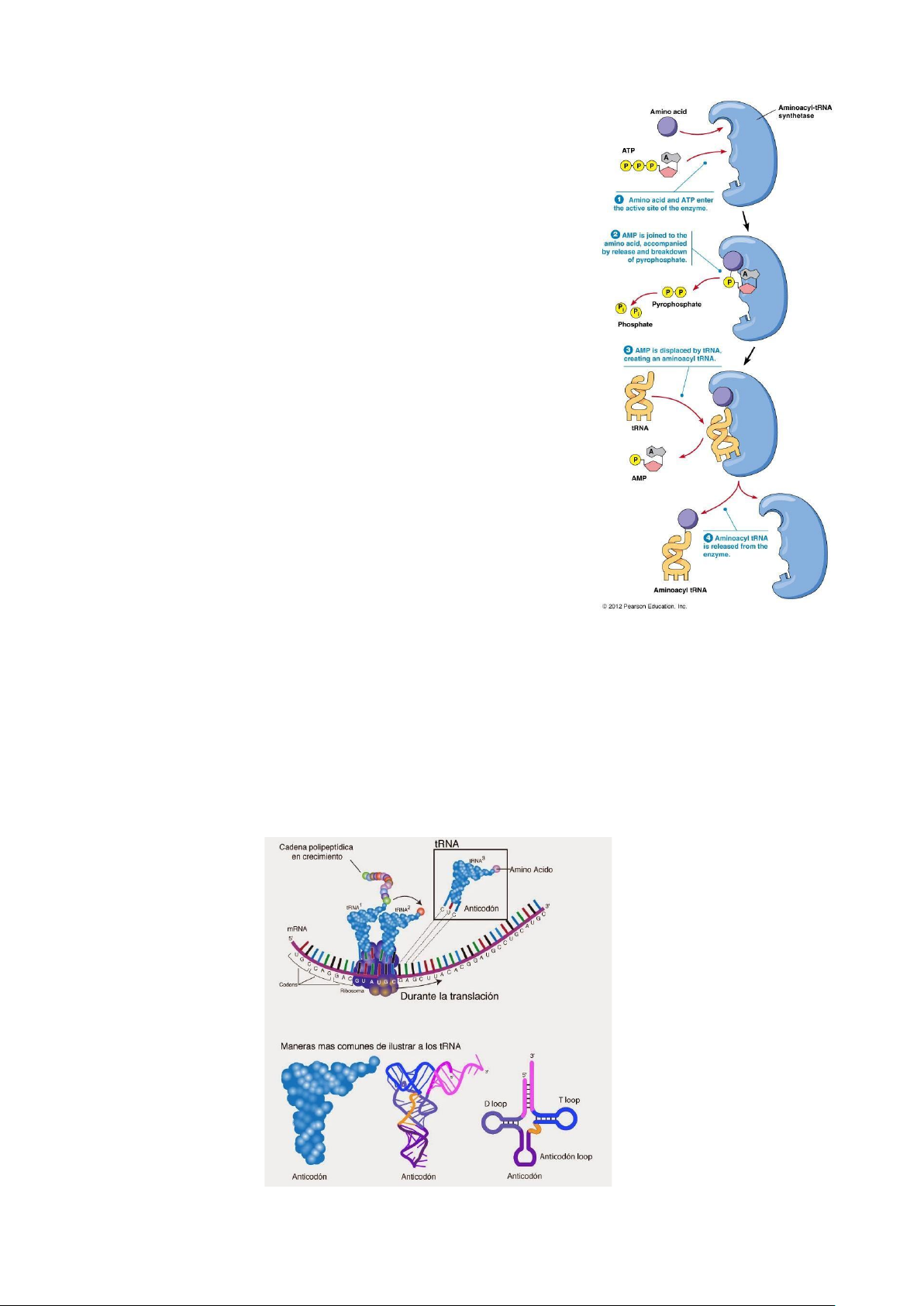
c. Quá trình tARN gắn acid amin:
- Là quá trình gồm 2 giai đoạn:
GĐ 1:Acid amin + ATP → Aminoacyl-AMP + PPi
GĐ 2:Aminoacyl- AMP + tARN → AminoacyltARN + AMP
- Enzym aminoacyl tARN syntherase: xúc tác giai đoạn aminoacyl hóa.
+ Mỗi enzyme nhận diện 1 acid amin đặc hiệu và
tARn vận chuyển acid amin đó.
+ Enzym nhận diện tARN ở hai vị trí: cuốn D và vòng đối mã.
- Cấu trúc enzyme aminoacyl tARN syntherase: + Nhóm 1 :
• Vị trí nhận diện : cuống D và vòng đối mã.
• Xúc tác 2—OH của Adenin. + Nhóm 2 :
• Vị trí nhận diện : cuống D.
• Xúc tác 3—OH của Adenin.
c. Cấu trúc không gian tARN -
Bậc 1: Trình tự nucleotide 5’-3’. - Bậc 2: Hình là chẻ 3.
- Bậc 3: Cấu trúc không gian 3 chiều hình chữ L → gắn vào vị trí A,P của Ribosome. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
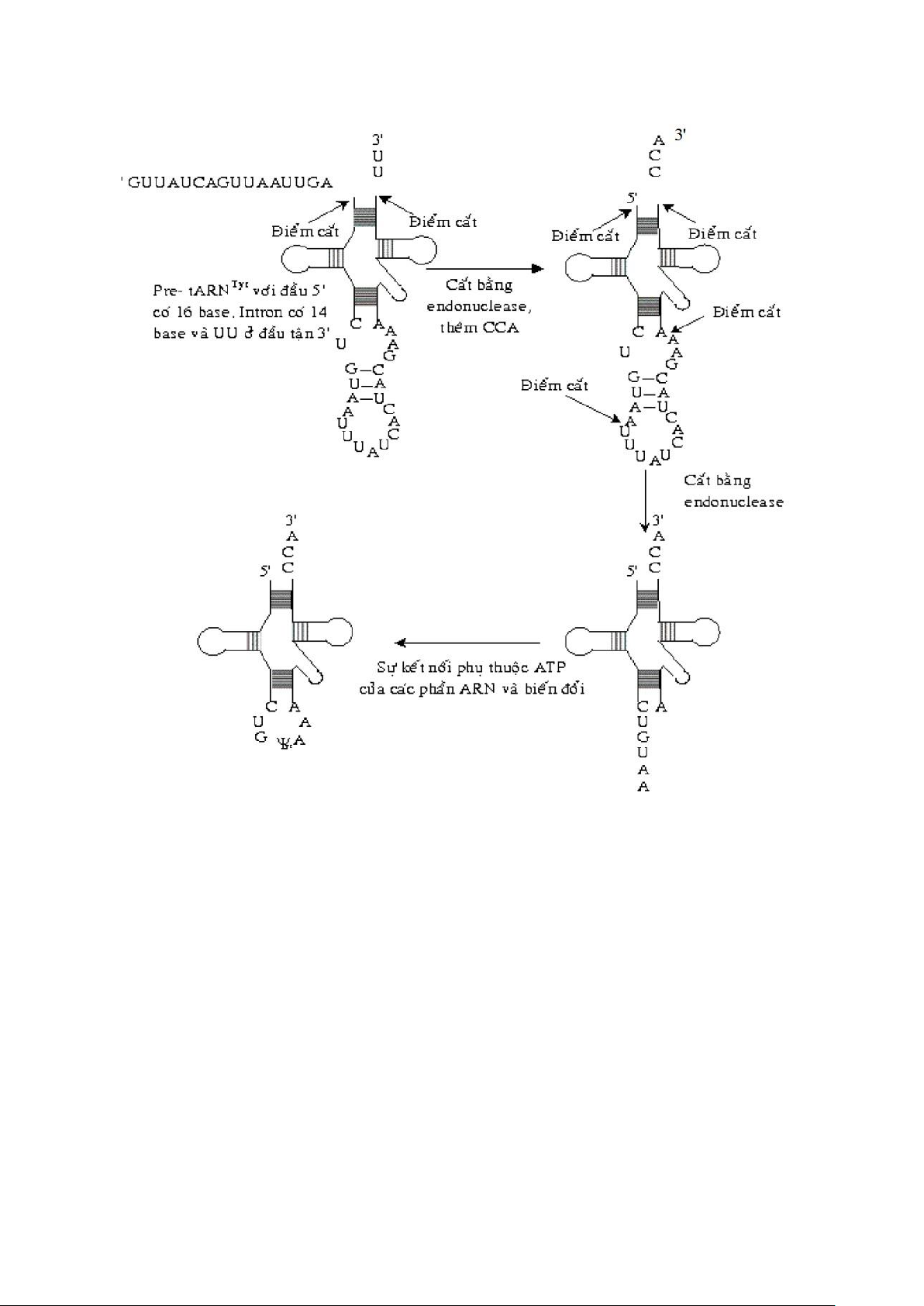
d. Quá trình biến đổi tARN.
- Biến đổi pre-tARN: + Loại bỏ intron vùng đối mã.
+ Cắt trình tự ở đầu 5’ bằng ARNase P( ARNase P được cấu tạo từ protein và ARN hoạt hóa bằng Mg2+.)
+ Trình tự ở đầu 3’ được cắt và thay thế bằng trình tự CCA. +
Biến đổi một số base.
- Đặc điểm quá trình biến đổi pre-tARN: + Intron không có trình ự nhận diện vị trí cắt. + Cần cung cấp GTP, ATP.
+ Vùng 2’,3’ – P được tạo thành ở exon 5’.
+ Cấu trúc bậc 2 của tARN ảnh hưởng đến qáu trình cắt nối. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ 3. mARN. a. Cấu trúc - Gồm 3 phần:
Đoạn 5’ không mã hóa - Vùng mã hóa - Đoạn 3’ không mã hóa.
=> Vùng mã không mã óa 5’UTR và 3’UTR có vai trò tăng hay giảm độ ổn
định ATN, giúp ARN tồn tại lâu hơn trong TB chất.
b. Xử lý hậu Phiên mã ở TB Eukaryote.
TB nhân thật, sau khi phiên mã → hnARN → xử lí hậu phiên mã → mARN
trưởng thành → Dịch mã. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
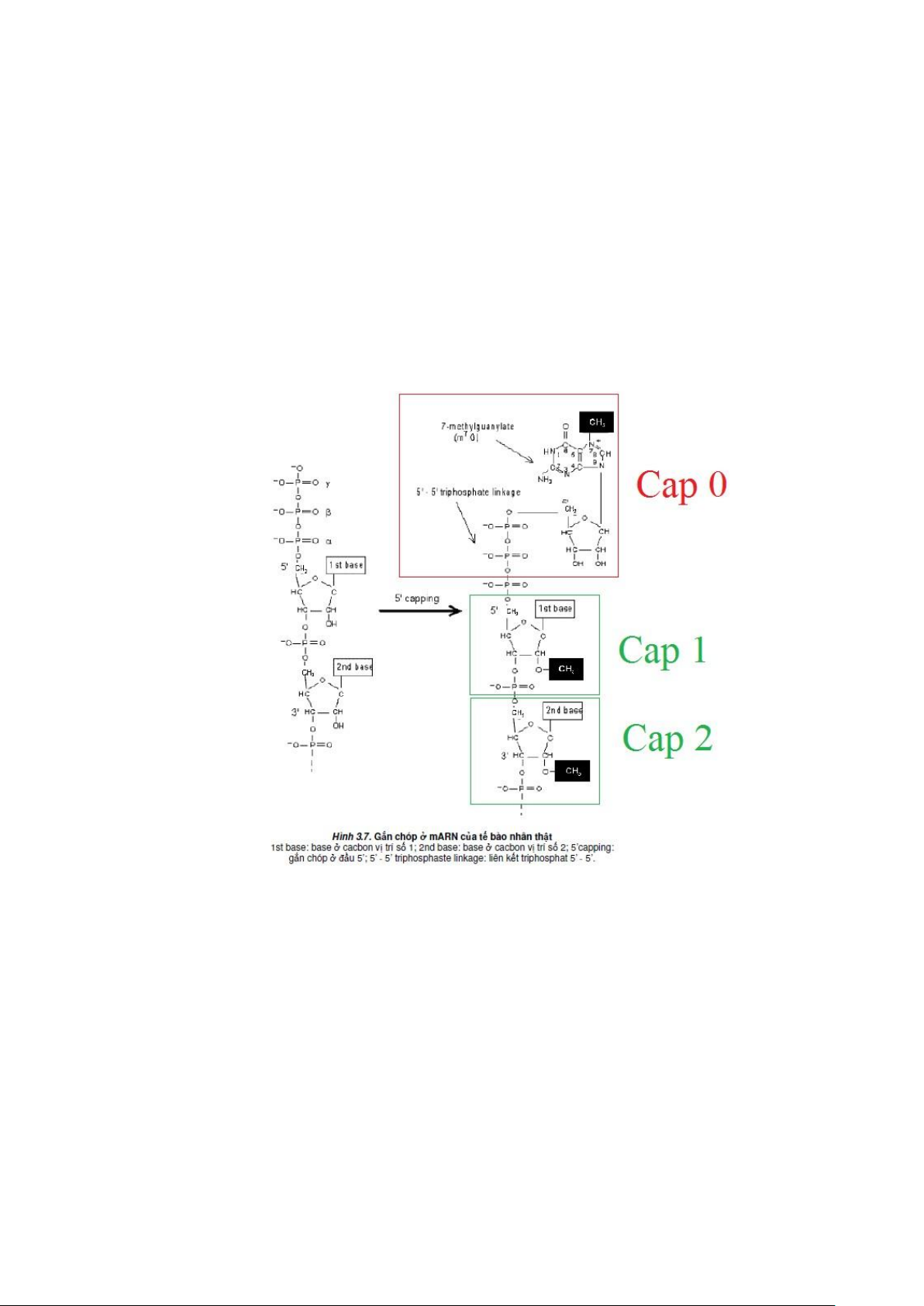
*Giai đoạn 1: Giai đoạn mũ hóa – gắn chóp.
- Gắn dạng đặc hiệu Guanin triphosphate và đầu 5’.
- Xảy ra theo một trong ba cách:
+ Cap O: Methyl gắn vào vị trí G7 do enzyme Guamin – 7 methyl transferase
xúc tác phản ứng xảy ra ở mọi mARN nhân thật.
+ Cap 1: Methyl gắn vào vị trí 2’ – OH do enzyme 2’ – O – methyl transferase
xúc tác phảm ứng xảy ra ở hầu hết mARn nhân thật.
+ Cap 2: Phản ứng methyl hóa tương tự xảy ra ở nucleotide thứ hau với tỉ lệ 10 – 15%.
*Giai đoạn 2: Giai đoạn gắn đuối PolyA vào 3’.
- Đuôi poly A ( 50 – 500 Adenin) gắn vào pre-ARN có trình tự nhận diện, do
enzyme poly A –polymerase xúc tác. - Vai trò đuôi Poly A:
+ Giúp mARN rời khỏi nhân.
+ Xác định số lần mARN được dịch mã. + Bảo vệ ARN. + Gắn với Ribosome.
Giai đoạn 3: Cắt nối:
- Cắt loại bỏ Intron, nối các đoạn Exon lại với nhau. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
c. mARN ngược.
- mARN có trình tự bổ sung với mARN → không thể dịch mã thành protein →
chống lại hiệu tượng gen nhãy ngược hay virus.
- Thêm mARN ngược → Nghiên cứu chức năng gen.
4. Các snARN và scARN.
a. snARN. snARN + Protein đặc hiệu → snRNP (ribonucleoprotein). b. scARN.
scARN + Protein đặc hiệu → scRNP (ribonucleoprotein). Nhận diện tín hiệu xuất.
III. Cắt nối ARN ở TB nhân thật.
1. Nguyên tắc GU – AU.
- GU luôn tận cùng đầu 5’ của intron vị trí cho.
- AG luôn tận cùng đầu 3 của intron vị trí nhận
→ bất kì một vị trí cho nào cũng có thể gắn với vị trí nhận.
2. Phản ứng cắt nối.
Hai phản ứng chuyển nhóm ester.
- Phản ứng chuyển nhóm Ester I: nhóm –OH ưa nhân gắn vào vị trí 5’P của intron.
- Phản ứng chuyển nhóm Ester II: nhóm 3’-OH ưa nhân của exon gắn vào vị trí cắt nối 3’. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
=> Nối các đoạn exon, cắt bỏ intron.
3. Cơ chế tự cắt nối ARN.
ARN đóng vai trò enzyme-ribozym. Có hai loại nhóm Intron nhóm I và Intron nhóm II.
4. Cắt nối nhờ Spliceosome.
- Splicesome là một phức hợp gồm khoảng 40 protein quy định cắt nối và các
snARN. Các phức ribonucleprotein này được gọi là snRNP hay snurp.
- Các snRNP gồm U1, U2, U5 và U4/U6 chứa các snARN U1,U2,U4,U5,U6.
Chúng nhận diện trình tự exon, intron để loại bỏ intron, nối exon.
- Quá trình cắt nối nhờ Sliceosome chỉ xảy ra ở SV nhân thật bậc cao. - Cơ chế:
+ Khi snRNP gắn vị trí cho của Intron, U4/U6 gắn với U2 → phức A chuyển thành phức B.
+ snRNP U1 phóng thích → snARN U6 tương tác vị trí cuối nối 5’ → chuyển phức B1 thành phức B2.
+ snRNP U4 tách khỏi snRNP U6 → snARN U6 bắt cặp với mARN U2 để tạo trung tâm xúc tác. NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
CHƯƠNG IV: PHIÊN MÃ VÀ MÃ DI TRUYỀN
I. Nguyên tắc chung Phiên mã.
- Chỉ một trong hai sợi ADN dùng làm mạch khuôn để tổng hợp ARN d9u7oc75
gọi là sợi khuông mẫu → Phiên mã là quá trình bất đồi xứng.
- ARN polymerase phụ thuộc ADN bám vào.
+ Gây đứt liên kết H giữa các cặp base bổ sung của chuỗi xoắn kép ADN →
tách thành 2 mạch đơn ADN.
+ Di chuyển 3’ – 5’ trên ADN → ARN được tổng hợp theo chiều 5’- 3’.
- Các nguyên tắc cảu phiên mã:
+ Cần cung cấp các ribonucleotide triphosphate: ATP, GTP, UTP, CTP.
+ ARN polymerase là 1 holoenzym → cần co-factor Mg2+ hoặc Mn2+
- Các yếu tố cần thiết cho phiên mã: + ADN khuông mẫu. + Co-factor: Mg2+, Mn2+. + ATP, GTP, UTP, CTP. + ARN polymerase.
- Enzym ARN-polymerase: không có hoạt tính sữa sai.
+ Các nucleotide kết hợp không chính xác có thể được thay thế ngay bằng
nucleoitide phù hợp. Có lỗi nặng → khó phiên mã.
+ Nếu có nucleotide gần sai → Không di truyền. - Phương trình phiên mã:
r(NMP)n + rNTP → r(NMP)n+1 + PPi
II. Quá trình phiên mã. 1. Quá trình chung.
- Khởi đầu → kéo dài → Kết thúc NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
- Sản phẫm phiên mã: ARN sơ khai → biến đổi → ARN trưởng thành. a. Phiên
mã bất đối xứng *Giai đoạn khởi đầu
- ARN-polymerase + yếu số sigma → bong bong phiên mã gắn với Promoter ADN.
*Giai đoạn kéo dài:
- ARN-polymerase cùng bong bong phiên mã chuyển dọc theo sợi ARN khuông
mẫu theo hương 3’ – 5’ → gắn các nucleotide trisphosphat.
- Khi phức hợp ADN-ARN đạt chiều dài 12 base → đầu 5’ của ARN tách khỏi
mạch đơn ADN bổ sung → Hai mạch kép của ADN ban đầu bắt đầu phục hồi.
*Giai đoạn kết thúc:
ARN- polymerase gặp điểm kết thúc → cắt đứt nối dài mạch ARN → ARN-
polymerase và chuỗi ARN hoàn chỉnh tách khỏi sợi ADN.
b. Phiên mã đối xứng
- Cả hai mạch ADN đều phiên mã thành ARN xảy ra ở ADN ty thể tế bào thú.
Nhưng say đó một trong hai ADN sẽ bị thoái hóa.
2. Phiên mã ở Prokaryote.
- Bộ NST của E.coli chứ khoảng 4.106 base pair, 3000 gen cấu trúc, 100 đơn vị
phiên mã. → có nhiều gen trong một đơn vị phiên mã.
*ARN polymerase ở E.Coli
+ Là phức hợp protein chứa 5 tiểu đơn vị thuộc 4 loại: α;β;β’;σ NGUYỄN ĐẮC NHÂN – D16 lOMoAR cPSD| 45470709 SINH HỌC PHÂN TỬ
+ Tiểu đơn vị σcó thể tách ra thuận nghịch từ holoenzym. α2ββ’σ → α2ββ’ + σ
(holoenzym) (core enzyme- lõi) (yếu tố tách ra)
• β và β’: chịu trách nhiệm xúc tác.
• α : liên quan đến lắp ghép enzyme.
• σ : liên quan đến tính dặc hiệu của promoter.
- Phiên mã không cần mồi do có vùng Promoter nhận diện bằng σ.
*Promoter phiên mã:
- Ký hiệu: Base đầu tiên Phiên mã trên mARN ký hiệu là +1 → các base ngược
chiều phiên mã ký hiệu “ – ”.
- Hai trình tự đặc biệt của Promoter phiên mã:
+ Hộp TATAAT ( Hộp TATA, hộp Pribnow):
• Thường tận cùng là base T.
• Cách điểm xuất phát 6-8 base.
• Base trung tâm ở vị trí âm 10. + Hộp TTGACA
• Base trung tâm ở vị trí âm 35.
• Tham gia gắn khởi đầu ARN-polymerase.
→ Không phải Promoter nào cũng có 2 trình tự trên có thể khác nhau vài nucleotide.
=> Hai trình tự trên của promoter cho phép gắn ARN-polymerase vào vùng khởi
động phiên mã. - Nhờ Đột biến điểm giúp xác đinh chức năng promoter.
*Promoter sock nhiệt: - Thí nghiệm:
+ Giả thuyết cho rằng ARN-polymerase có một holoenzym phiên mã tất cả các gen.
+ Thí nghiệm: gây gián đoạn nhiệt môi trường nuôi cấy
+ Kết luận: khả năng phiên mã do tiểu đơn vị σ khác nhau → chỉ có protein
sock nhiệt mới giúp Tế bào chống lại sự biến đổi nhiệt. → Chỉ có
ARNpolymerase σ32 mới có khả năng gắn với Promoter sock nhiệt phiên mã ra
ADN sock nhiệt. NGUYỄN ĐẮC NHÂN – D16
Tài liệu liên quan:
-

Bài giảng sinh học phân tử | Đại học Y Dược Thành phố Hồ Chí Minh
55 28 -

Bài tập sinh di truyền cho môn sinh học phân tử | Đại học Y Dược Thành phố Hồ Chí Minh
60 30 -

Bài 6: Điều Hòa Hoạt Động Gen - Câu Hỏi Sinh Học Phân Tử | Đại học Y Dược Thành phố Hồ Chí Minh
60 30 -

Bài 3 - Các loại ARN: Câu hỏi trắc nghiệm Sinh học phân tử | Đại học Y Dược Thành phố Hồ Chí Minh
61 31 -

Câu hỏi ngắn Sinh học phân tử | Đại học Y Dược Thành phố Hồ Chí Minh
110 55