Đề cương sinh học phân tử –Trường Đại học Nguyễn Tất Thành
+ Giai đoạn SHPT ra đời
1953 Watson, Crick: xác định mô hình cấu trúc xoắn kép DNA
1958 Francis: học thuyết trung tâm – Central Dogma “ thông tin di truyền một khi đã chuyển sang protein thì không thể lấy lại được”
1961 M. Nirenberg và J. Matthei: tìm ra bộ mã di truyền
Đề cương sinh học phân tử –Trường Đại học Nguyễn Tất Thành
Tài liệu gồm 16 trang , giúp bạn tham khảo, ôn tập và đạt kết quả cao
Môn: Sinh học phân tử 28 tài liệu
Trường: Trường Đại học Nguyễn Tất Thành 1.4 K tài liệu
Tác giả:











Preview text:
BÀI 1: NHẬP MÔN SINH HỌC PHÂN TỬ -
Giai đoạn hình thành và phát triển SHPT: hình thành tiền đề
-> SHPT ra đời -> SHPT hiện đại
+ Giai đoạn tiền đề
1869 Miescher: phát hiện nuclein ( không phải protein) -> acid nucleid
1865 Medel: quy luật di truyền tính trạng trên đậu Hà Lan
1928 GrifÏth: chuyển đổi kiểu hình R (không gây bệnh) sang
S (gây bệnh) ngay cả khi dùng tb chết
1944 Avery, MacLeod, McCarty: công bố bằng chứng DNA là vật liệu di truyền
1952 Hershey và Chase: xác nhận DNA là vật liệu di truyền bằng kỹ thuật phóng xạ
1951 E. Chargaff: chứng minh trong DNA tỷ lệ base purin bằng tỷ lệ
base pyrimidin ( A=T, G=C); tỷ lệ A+T khác G+C và thay đổi theo loài
+ Giai đoạn SHPT ra đời
1953 Watson, Crick: xác định mô hình cấu trúc xoắn kép DNA
1958 Francis: học thuyết trung tâm – Central Dogma “ thông tin di
truyền một khi đã chuyển sang protein thì không thể lấy lại được”
1961 M. Nirenberg và J. Matthei: tìm ra bộ mã di truyền đầu tiên
1966 cùng H.G Korana: giải mã toàn bộ 64 codon
1961 F. Jacob và J. Mono: cơ chế điều hòa tổng hợp protein
1962 W. Aber, D. Nathans, H. Smith: tìm ra enzym cắt giới hạn
1967: chiết xuất được DNA ligase
+ Giai đoạn SHPT hiện đại
Thập niên 70 TK XX: xác định trình tự nucleotid trên gen -> gây đột
biến định hướng cho các biến đổi tùy ý
1990: nghiên cứu insillico ( trên máy điện toán ) tạo thuận lợi cho nghiên cứu sinh học
1970 – H. Smith: chiết được enzyme cắt giới hạn
1972 – Paul Berg: tạo ra DNA tái tổ hợp trong ống nghiệm
1973 – A.C. Chang, H. Boyer, S. Cohen … tạo ra plasmid
tái tổ hợp, ứng dụng E.coli → thúc đẩy sự ra đời của công nghệ di truyền
1977 W. Gilbert và F. Sanger: tìm ra kỹ thuật xác định trình tự DNA
1985 R. K. Saiki và K. B, Mullis : kỹ thuật PCR -> ứng dụng trong
chẩn đoán, biến đổi di truyền, xác định phả hệ,…
Khái niệm: SHPT là một bộ phận của Sinh học, khoa học về sự sống
Đối tượng nghiên cứu: sự sống cấp độ phân tử
Tập trung các khía cạnh về cấu trúc
Sao chép về biểu hiện của gen
Tương tác và chức năng sinh lý của sản phẩm gen
Phương pháp nghiên cứu
Tạo dòng biểu hiện gen: cô lập gen để nghiên cứu biểu hiện gen
PCR: khuếch đại gen đưa đột biến vào
Điện đi gel: phân tách gen, protein
Lai vết Southern DNA Northen:phát hiện gen, nghiên cứu sự biểu hiện
Lai vết Western/ hóa miễn dịch: phát hiện protein, nghiên cứu chức năng protein
Đóng góp của SHPT Genomic: giải mã bộ gen
Proteomics: phan tích biến động protein Sự phát triển thuốc Chip DNA
Chuyển gen vào cây trồng Tin sinh học Công nghệ nano sinh học
BÀI 2 SAO CHÉP DNA
Là sự khởi đầu cần thiết cho sự sinh sản
DNA hoặc RNA ( virus) là nơi cất giữ thông tin di truyền
TB phân chia -> DNA phải được sao chép -> đảm bảo thông tin
di truyền chuyển cho TB con
Yếu tố vật lý, hóa học,… có thể tác động lên DNA -> phá huy DNA ->
sửa chữa, làm chết TB, biến đổi gen -. Di truyền sang thế hệ sau
Cấu trúc Deoxyribonucleic Acid
Mạch đôi, xoắn kép: 2 chuỗi polynicleotid xoắn quanh 1 trục theo hướng ngược nhau Đường Deoxyribose ( 5C) Gốc phosphat
Các base nito Adenin, Guanin, Cytosin và Thymin
SAO CHÉP DNA
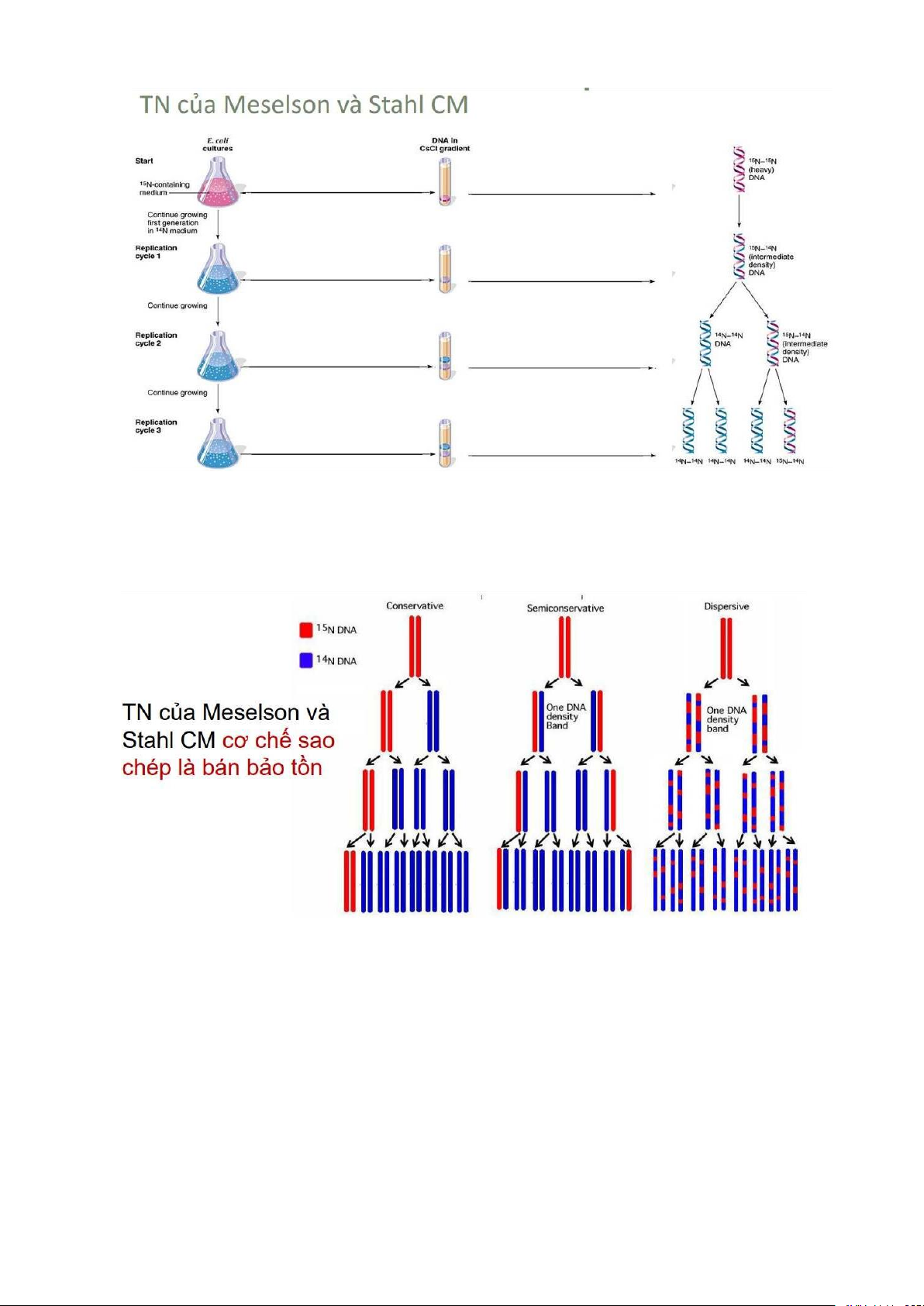
Mô hình sao chép
Bảo tồn: phân tử ADN con - gồm 2 chuỗi hoàn toàn mới
Bán bảo tồn: phân tử ADN con: gồm 1 chuỗi mẹ kết hợp với 1
chuỗi mới được tổng hợp
Phân tán: phân tử DNA con chứa các đoạn rải rác của DNA mẹ -
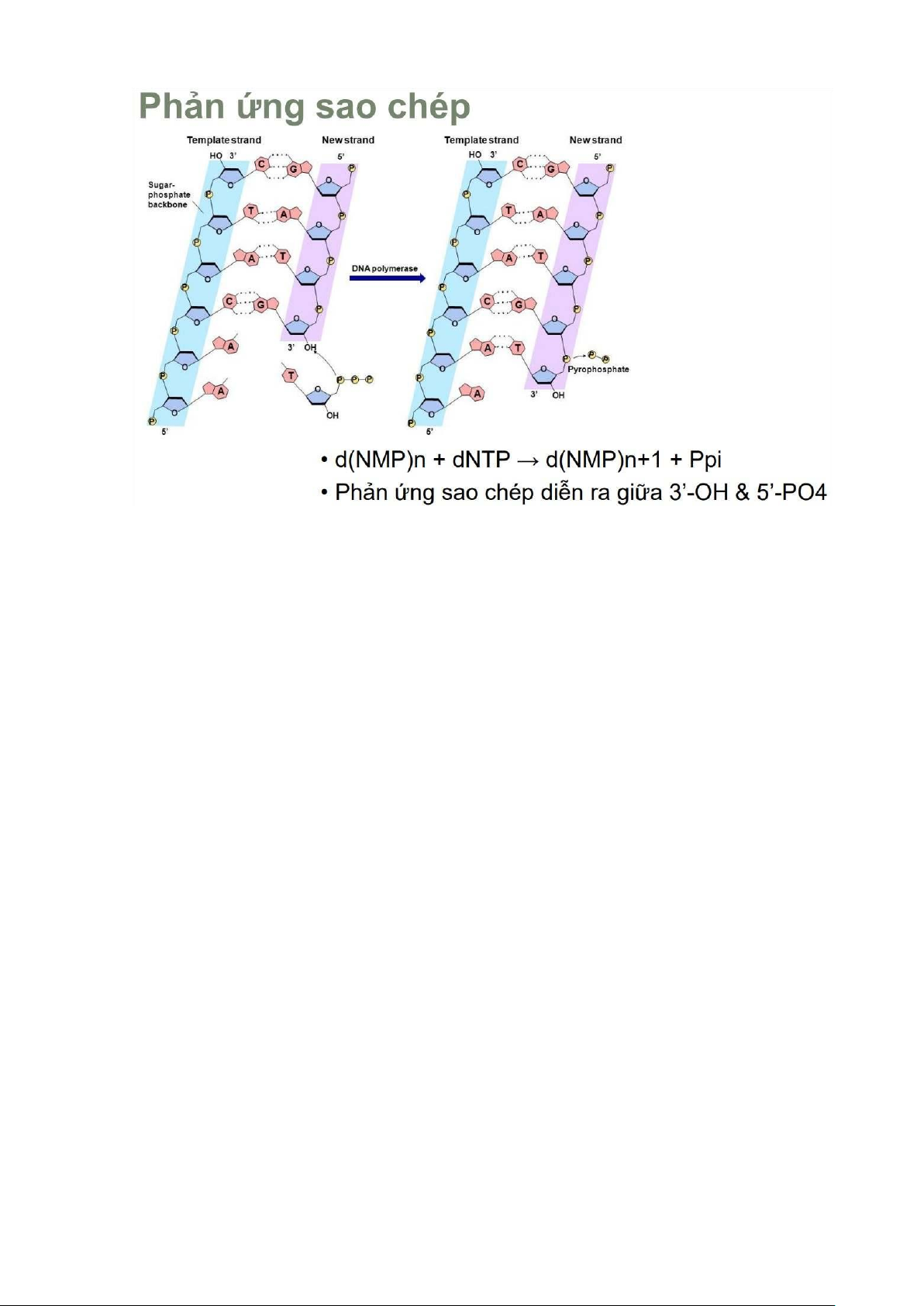
Phản ứng sao chép •
Mạch DNA được kéo dài nhờ sự thành lập liên kết
phosphodiester giữa mạch cũ và nucleotid mới •
Bazo nito lk góc đường ở C1 •
Gốc P lk góc đường ở C5 •
Gốc OH lk gốc đường ở C3 -
Yếu tố cần thiết cho sao chép DNA: khuôn mẫu, Mg2+, 4
loại deoxyribosenucleotid triphosphat (dNTP: A, T, G, C), enzyme DNA
polymerase, Primase, RNAse H, DNA ligase, SSB protien – singel
strDNA binding protein, heliacse, Topoisomerase ( I,II) -
DNA Polymerase: phụ thuộc DNA
+ Xúc tác sự thành lập liên kết phosphodiester: polymer hóa theo
hướng 5’ – 3’ -> kéo dài chuỗi DNA •
Exonuclease 5’ - 3’: hủy mồi của ADN pol I •
Exonuclease 3’ - 5’: sửa lỗi
+ E. coli: ADN polymerase I, II, III, IV, V + Nhân thật: ADN polymerase α, β, γ, δ và ε - SAO CHÉP
DNA Ở E. COLI: khởi đầu ( tạo chạc ba sao
chép) -> kéo dài ( liên tục trên sợi sớm, không liên tục trên sợi muộn) - > kết thúc
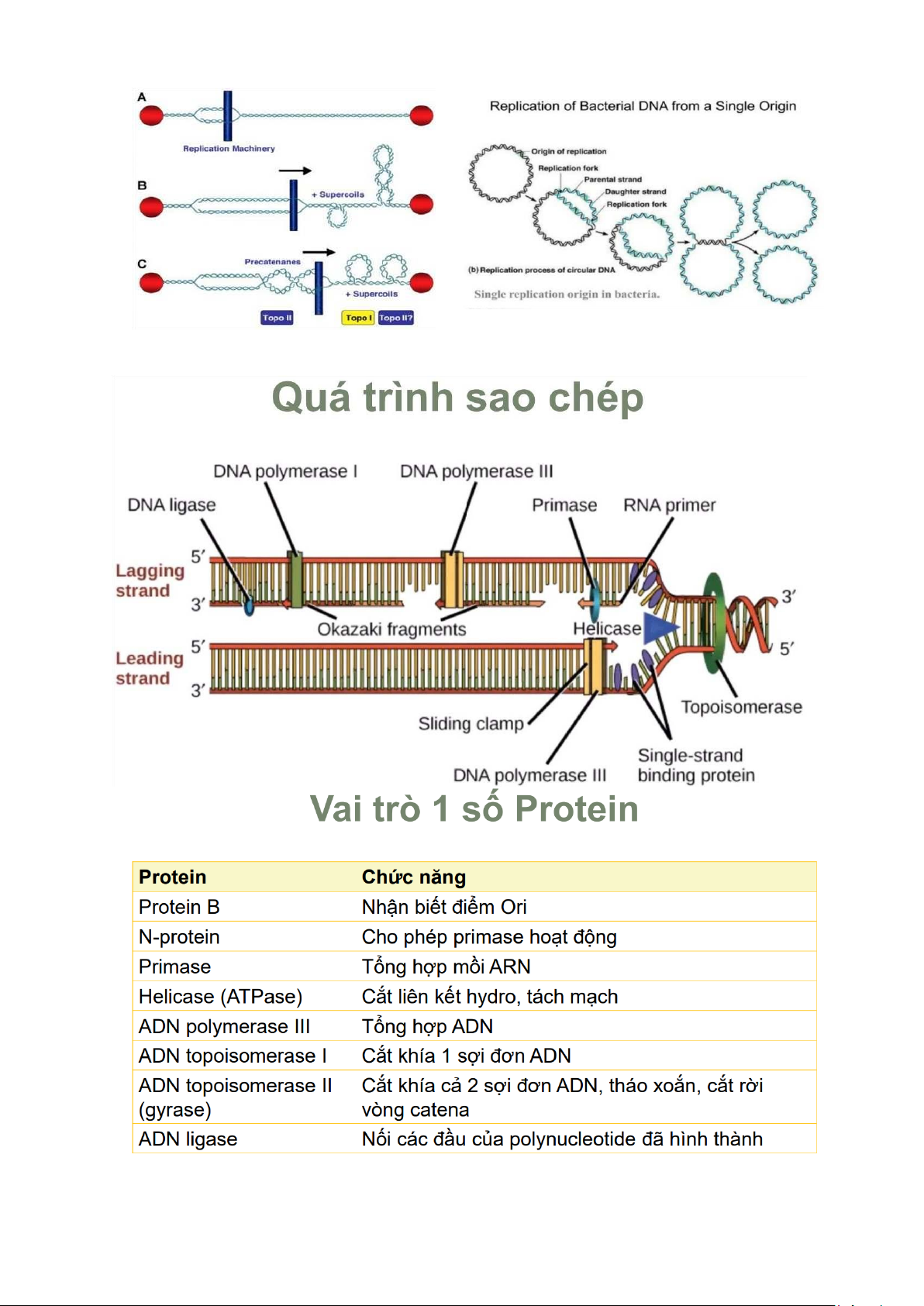
+ Khởi đầu sao chép •
Protein B nhận biết điểm khởi đầu sao chép ( Ori C) giàu A – T ( 254 cặp base) •
Gyrase (topoisomerase II): tháo xoắn AND •
Helicase (ATP: cofactor): cắt lk hydro, tách mạch •
SSB protein gắn vào, giữ sợi đơn không chập lại ⇨
Hình thành chạc ba sao chép
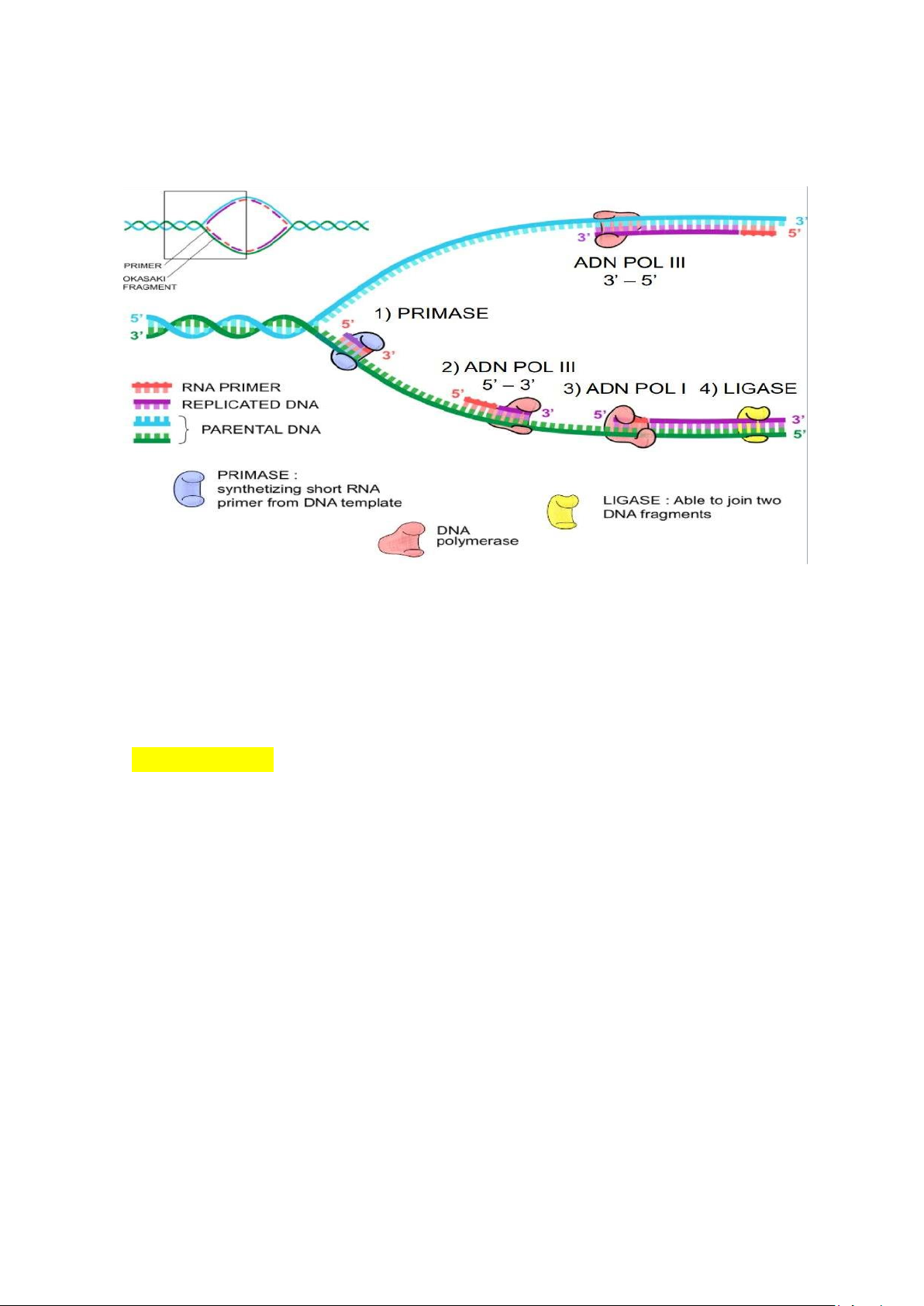
+ Kéo dài sao chép •
Enzyme Primase tạo mồi ( nhiều vị trí trên sợi muộn) •
Sao chép trên sợi sớm
Sợi sớm: sợi con bổ sung với mạch khuôn 3’ - 5’ Hướng sao chép 5’ - 3’: sao chép liên tục
ADN polymerase III: gắn vào mạch khuôn (3’ - 5’), lắp nucleotid bổ sung và kéo dài mạch •
Sao chép trên sợi muộn
Sợi muộn: sợi con bổ sung với mạch khuôn ADN 5’ - 3’ Hướng sao
chép 5’-3’: sao chép ko liên tục
ADN pol III gắn vào, kéo dài mồi theo hướng 5’-3’ tạo các đoạn Okazaki (1000-2000 nu)
ADN pol I cắt bỏ mồi (exonuclease 5’ – 3’), lấp đầy các nucleotid ADN vào chỗ trống
Ligase nối các đoạn Okazaki lại
+ Kết thúc sao chép •
Sự kéo dài được giáp vòng hay đến đầu mút của AND •
Kết thúc sao chép, 2 sợi đôi ADN lồng vào nhau → vòng lồng ghép ⇨
Cần topoisomerase II (gyrase) để tách 2 ADN vòng bị lồng vào nhau (catena)
# CẤU TRÚC THETA
Giải pháp ngăn chặn siêu xoắn •
Topoisomerase I: cắt khía 1 sợi DNA, DNA quay quanh sợi
còn lại -> mất xoắn ; không cần ATP •
Topoisomerase II: cắt 2 sợi DNA -> mất vòng lồng ghép ; cần ATP -
SAO CHÉP DNA Ở TẾ BÀO NHÂN THỰC •
ADN TBNT có nhiều replicon: đơn vị sao chép •
Saccharomyces cerevisiae có 500 replicon, tức 500 điểm Ori •
Cơ chế tương tự tế bào nhân nguyên thủy
Polymerase δ tổng hợp sợi sớm Polymerase α tổng hợp sợi muộn
Polymerase β sửa sai
Sao chép ADN ở TBNT khác nhân sơ: •
Tốc độ di chuyển ADN polymerase chậm hơn: do ADN
đóng cuộn trong NST và dài hơn •
Tốc độ sao chép nhanh hơn: nhiều enzyme và điểm replicon (Ori) •
Các đoạn Okazaki nhỏ hơn, khoảng 40-300 base -
SAO CHÉP DNA TY THỂ •
Enzym sao chép: ADN polymerase γ •
Bắt đầu ở 1 sợi ADN gốc •
Sự sao chép tiến hành chưa được ½ đường → bắt đầu sao chép
sợi còn lại tạo cấu trúc chữ D •
Topoisomerase II: tách đôi 2 vòng kép ADN ty thể I.
CÁC KIỂU SAO CHÉP (2 kiểu) - KIỂU THETA •
Sao chép xảy ra theo 2 chiều cùng lúc tại 1 điểm Ori • Sản phẩm DNA mạch vòng -
KIÊU LĂN VÒNG •
1 trong 2 mạch làm khuôn, giữ nguyên cấu trúc & xoay trong quá trình sao chép •
Sản phẩm DNA mạch thẳng
??? Sao chép kiểu dạng thẳng -> bộ gen ngắn hơn sau mỗi lần sao chép Cách giải quyết •
Phage T7: thành lập phức nối •
Phage λ vòng hóa bộ gen nhờ trình tự cos -
Sao chép DNA ở Phage T7 •
Bộ gen là ADN sợi đôi, thẳng •
Trình tự Nu ở đầu 3’OH của 2 sợi bổ sung cho nhau •
Trình tự cos: 160 bp đầu tiên bên trái (3’ -OH) = 160 bp cuối cùng bên phải (3’ - OH) -
Sao chép DNA ở Phage LAMDA •
Trình tự cos: 160 bp đầu tiên bên trái (3’ -OH) = 160 bp
cuối cùng bên phải (3’ - OH) •
Vị trí cos ở 2 đầu mút bộ gen dài 12 Nu: bộ gen thẳng vòng đôi •
Khi xâm nhiễm E.coli, phage bơm ADN vào, vòng hóa bộ gen
tránh bị cắt bởi exonuclease •
Gồm 2 giai đoạn :
GĐ sớm (kéo dài ~ 15’): sao chép theo mô hình theta
GĐ muộn: sao chép theo kiểu lăn vòng
→ Sản phẩm: ADN thằng kéo dài chứa nhiều bản sao liên tiếp, cách
nhau bằng các trình tự cos
→ Terminase: nhận diện trình tự cos, cắt ADN dài thành nhiều bản sao đơn của phage -
sao chép ở Phage 13 •
Bộ gen: ADN sợi đơn, vòng •
Phụ thuộc bộ máy sinh tổng hợp của E.coli •
Khi xâm nhiễm E.coli, bơm ADN vào & bao bọc bởi SSB protein •
Tổng hợp ADN sợi đôi: thể sao chép RF1 nhờ cấu trúc kẹp tóc sợi đôi trên ADN phage •
Sao chép theo kiểu lăn vòng nhờ gp 2 endonuclease •
Sản phẩm: ssADN vòng (sợi mới tạo ra được nối thành vòng), ADN sợi đôi RF II.
QUÁ TRÌNH SỬA SAI -
Trong sao chép •
Tần suất sai sót invitro: 10 mũ trừ 5; invivo: 10 mũ trừ 9 •
Nhân nguyên thủy: ADN polymerase I và III vừa polymer hóa
vừa sửa sai nhờ hoạt tính exonuclease 3’ – 5’ •
Nhân thật: ADN polymerase δ và ϵ; ADN polymerase α
KHÔNG có hoạt tính sửa sai •
Ty thể: polymerase γ không có hoạt tính exonuclease -
Khi không sao chép •
Enzym chuyên biệt ( khoảng 50 enzyme) dò tìm và sửa sai
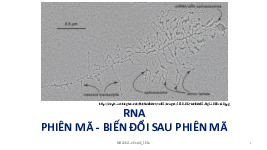
BÀI 3: CẤU TRÚC RNA I. RNA -
Thành phần: A, U, G, C
# Khác với DNA:
RNA: 1 sợi đơn, đường ribose, thành phần (A,U,G,C)
# nucleotid: phosphate, đường ribose, nitrogen base (A,U,G,C) -
mRNA – messenger RNA
+ vị trí: bắt đầu ở nhân và di chuyển đến ribosome
+ chức năng: mang thông tin mã hóa amino acid
+ cấu trúc 3 phần: 2 vùng không mã hóa (tồn tại lâu -> tổng hợp
nhiều protein); 1 vùng mã hóa
# sinh ra ở nhân -> mang thông tin di truyền vào ribosome ->
ribosome được tổng hợp (dịch mã)
+ sự biến đổi mRNA ở TBNT •
giai đoạn găn chóp đầu 5’ ( capping): chóp này dùng bảo
vệ RNA; làm vị trí để ribosome nhận diện gắn vào và dịch mã) •
giai đoạn gắn đuôi ở đầu 3’ ( poly A) + quá trình cắt nối
• mRNA nguyên thủy gồm exon và intron
⇨ Exon mang mã di truyền của protein ⇨
Intron ( lớn hơn exon) xen kẽ giữa các exon • Cắt intron – nối exon •
Xúc tác bởi RNA : snRNP (ribonucleoprotein nhân nhỏ) =
snRNA ( RNA nhân nhỏ) + protein
• Nguyên tắc cắt nối pre – mRNA : GU – AG
⇨ GU luôn ở tận cùng 5’ intron: vị trí cho
⇨ AG luôn tận cùng 3’ intron: vị trí nhận - tRNA – transfer RNA
+ cấu trúc : chẻ ba bền vững, chứa anticodon, có đuôi 3’ CCA để gắn với amino acid
+ vị trí: tế bào chất
+ chức năng : vận chuyển amino acid đến ribosome-> tổng hợp protein
# ribosome nối các amino acid ( dịch mã)
+ cơ chế hoạt động: enzym kết hợp amino aicd -> có mặt ATP nên hoạt
động -> phức hợp bắt tRNA -> tRNA được gắn amino aicd •
Việc gắn amino acid vào tRNA đặc hiệu được thực hiện qua
trung gian của enzyme amino acyl-tRNA synthetase •
Mỗi tRNA chỉ vận chuyển 1 loại amino acid xác định •
Mỗi amino acid được vận chuyển bới 1 số tRNA khác nhau (codon đồng nghĩa) •
Quá trình gắn amino acid vào tRNA gồm 2 giai đoạn: ⇨
Giai đoạn 1: enzyme nhận biết và gắn với 1 amino acyl đặc hiệu
Enzyme + amino acid +ATP → Enzyme-aminoacyl-AMP + P- P ⇨
Giai đoạn 2: Amino acid chuyển từ phức hợp enzyme-
aminoacyl sang tRNA tương ứng
Enzyme-aminoacyl-AMP + tRNA → tRNA-aminoacyl + AMP + Enzyme -
rRNA – ribosome RNA
+ tùy theo hệ số lắng S ( sendimention) trong quá trình ly tâm
phân tích, rRNA được chia thành nhiều loại khác nhau
+ vị trí: sản xuất trong nhân -> di chuyển ra tế bào chất
+ cấu trúc: hình thành bởi nhiều loại ribosome khác nhau; 2 tiểu đơn
vị tách ra hoặc gộp lại một cách thuận nghịch tùy theo Mg2+ # tiểu
đơn vị : rRNA + protein
# ribosome : ở vi khuẩn và lục lạp 70S; TBNT 80S; ty thể 50S
+ chức năng: gắn với các protein đặc biệt để tạo thành ribosome, một
số có khả năng tự cắt (rybozyme)
+ tách ra: Mg2+ trong môi trường thấp
+ gộp lại: Mg2+ trong môi trường cao -> tham gia dịch mã
+ các loại RNA •
RNA nhân không đồng nhất (hnRNA – heterogeneous
nuclear hay pre – mRNA) ⇨ Chỉ thấy ở TBNT ⇨
Tiền nhân của mRNA, được hình thành trong quá trình xử lý sau phiên mã •
RNA nhân nhỏ ( snRNA) ⇨ Chỉ thấy ở TBNT ⇨
Chức năng chính là tham gia vào cắt nối ( loại bỏ intron) mRNA
ScRNA: RNA tế bào chất nhỏ
Pre -rRNA: tiền rRNA Pre – tRNA: tiền tRNA
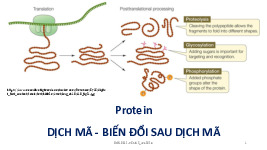
Quá trình phiên mã ( Transcription)
Đặc điểm cơ bản của quá trình phiên mã -
chỉ dùng 1 mạch để làm khuôn - không cần mồi (primer) -
RNA polymerase tháo xoắn (3’ – 5’) và tổng hợp mạch (5’ – 3’) -
Chỉ chọn lọc một số đoạn trên genome -
RNA polymerase không có khả năng sửa sai (tuổi thọ RNA ngắn,
nếu phiên mã sai -> không di truyền) -
Cơ chất: nucleotide triphosphate (ATP, GTP, UTP,CTP) -
Cả 3 loại RNA được tổng hợp từ DNA
- Phiên mã ở Prokaryote
+ σ: Nhận diện promotor
+ β’ : liên kết với sợi DNA khuôn
+ β: hình thành liên kết phosphodiester
+ Vị trí khởi động (promoter): trình tự gồm 6 nucleotide, cách điểm
bắt đầu tổng hợp RNA 10bp (trình tự -10: TATAT) và cách điểm bắt đầu
tổng hợp 35bp (trình tự -35: TTGACA)
+ Giai đoạn mở đầu •
Quá trình tổng hợp RNA được tiến hành khi RNA-
polymerase nhận ra và bám chặt vào vùng promotor nhờ yếu tố σ tạo
thành một phức hệ promotor mở •
Tiểu đơn vị σ giúp RNA polymerase nhận biết và gắn vào vị trí
trình tự -35, làm cho trình tự -1 tháo xoắn, từ đó một sợi đơn DNA được
sử dụng làm khuôn để tổng hợp RNA
+ Giai đoạn kéo dài •
NTP đầu tiên được nối, hình thành cặp base tại +1 •
ARN di chuyển dọc AND theo chiều 3’-5’, kéo dài mạch ARN nhờ lk phosphodiester •
Kéo dài 12 Nu: σ tách ra, đầu 5’ mARN tách khỏi AND
+ Giai đoạn kết thúc •
Trên DNA của vi khuẩn có các dấu hiệu kết thúc, khi RNA
polymerase gặp dấu hiệu này sẽ ngừng phiên mã, giải phóng RNA và
xúc tác cho quá trình phiên mã mới. Phân tử tRNA và rRNA sau khi giải
phóng sẽ tiếp tục hoàn thiện cấu trúc bậc 2 -
Cấu trúc kẹp tóc: trên sợi RNA có trình tự lặp và bổ sung ->
bắt cặp tạo thành hình dạng kẹp tóc -> làm cho RNA polymerase không
di chuyển được -> dừng lại
Phụ thuộc Rho: Rho cắt lk RNA ⇨
Trên DNA mang các trình tự báo hiệu chấm dứt hoạt động
phiên mã, được gọi là dấu hiệu kết thúc ⇨
Rho là một protein gồm 6 tiểu đơn vị giống nhau, ái lực cao với RNA ⇨
Rho nhận diện một vùng trên RNA gọi là “rut” gồm 50 –
90bp nằm trước trình tự kết thúc, giàu C và ít G ⇨
Rho có hoạt tính như enzyme helicase sẽ tách cấu trúc kép
RNA-DNA, giải phóng DNA và chấm dứt quá trình phiên mã
Không phụ thuộc Rho: gặp kẹp tóc -> dừng lại ⇨
Trên DNA mang các đoạn lặp đảo ngược tạo cấu trúc kẹp tóc ⇨
Sau cấu trúc kẹp tóc: 6 nucleotide adenine được PM thành chuỗi polyuracyl ⇨
Cấu trúc kẹp tóc sẽ làm cho RNA polymerase di chuyển
chậm và dừng lại dẫn đến chấm dứt quá trình phiên mã.
- Phiên mã ở Eukaryote + xảy ra trong nhân
+ nhiều loại RNA polymerase
+ tương tự TBNT nhưng phức tạp hơn: preRNA phải qua biến đổi và
dịch mã. Thường gồm 2 quá trình chính •
Quá trình phiên mã tạo nên phân tử mRNA đầu tiên, gọi là pre- mRNA •
Quá trình biến đổi phân tử pre-mRNA thành phân tử mRNA trưởng thành
+ dị và nguyên nhiễm sắc
+ cấu trúc gene ở Eukaryote •
Vùng 5’ kiểm soát biểu hiện gen: vùng này gồm các trình
tự nucleotide điều hòa biểu hiện gene và hoạt hóa sự phiên mã •
Vùng được phiên mã: ⇨
Gồm các exon và intron nằm xen kẽ. Đây là đặc điểm phân biệt với prokaryote ⇨
Các exon và intron đều được phiên mã nhưng chỉ có exon
mang mã di truyền được dịch mã ⇨
Các intron được bắt đầu bằng GT và kết thúc bằng AG ⇨
Intron chiếm phần lớn trong mỗi gene và sẽ được loại bỏ
khỏi RNA mới được tổng hợp, còn các exon được nối với nhau tạo thành mRNA hoàn chỉnh •
Vùng 5’ không dịch mã: tính từ vị trí bắt đầu phiên mã đến cocon khởi đầu ATG •
Vùng 3’ không dịch mã: bắt đầu từ codon kết thúc đến vị trí gắn đuôi poly (A)
+ giai đoạn khởi đầu: TFIID có chuỗi peptid TBP gắn với TATA box (
định hướng phiên mã) -> bẻ cong DNA tạo thành quai -> gắn với vùng
tăng cường -> điều khiển quá trình phiên mã
# yếu tố phiên mã có bản chất là protein •
Vai trò RNA polymerase I : RNA polymerase I cần có 2
protein là SL1 và UBF gắn vào 2 vùng chức năng giàu GC và có trình tự
giống nhau trên DNA (Vị trí -40 đến +20 core element ; Vị trí -180 đến -107 control element) •
Vai trò RNA polymerase III : Chỉ xúc tác phiên mã các
RNA nhỏ như tRNA, RNA 5S, snRNA
+ giai đoạn kéo dài : mRNA được tổng hợp trên mạch khuôn của DNA
+ giai đoạn kết thúc : •
RNA polymerase I: cần yếu tố kết thúc phiên mã giống
như Rho ở vi khuẩn, yếu tố này gắn với 1 trình tự DNA sau vị trí kết thúc •
RNA polymerase III: phiên mã một trình tự kết thúc tạo một
chuỗi uracyl trên phân tử RNA mà không cần hình thành cấu trúc kẹp tóc như ở vi khuẩn •
RNA polymerase II: kết thúc phiên mã tại nhiều vị trí trong
khoảng từ vài tram đến vài nghìn cặp base phía cuối gene
Vùng tăng cường: tăng cường quá trình dịch mã
#video: RNA polymerase bám vào một đoạn gen, tháo xoắn, tách mạch
-> phiên mã hoàn thành, mRNA được đưa ra ngoài -
Phiên mã ngược RETROVIRUS •
Phiên mã ngược là một quá trình sinh học đặc biệt, RNA
dung làm khuôn để tổng hợp DNA, đi ngược lại với nguyên lý "di
truyền một chiều" của sinh học phân tử. •
Quá trình này chủ yếu xuất hiện ở một số loại virus, đặc biệt là retrovirus như HIV •
Enzyme tham gia chính là enzyme reverse transcriptase
+ các bước: tạo DNA bổ sung ( cDNA) -> tạo cDNA sợi kép -> chèn
DNA vào bộ gen của tế bào chủ
+ ứng dụng: nghiên cứu và phát triển thuốc điều trị HIV; ứng dụng
trong công nghệ di truyền; công nghệ PCR phiên mã ngược (RT-PCR)
Tài liệu liên quan:
-
RNA phiên mã - biến đổi sau phiên mã môn sinh học phân tử | Trường Đại học Nguyễn Tất Thành
28 14 -
Protein dịch mã - biến đổi sau dịch mã môn sinh học phân tử | Trường Đại học Nguyễn Tất Thành
29 15 -
Đề cương sinh học phân tử –Trường Đại học Nguyễn Tất Thành
52 26 -

Tài liệu sinh học phân tử –Trường Đại học Nguyễn Tất Thành
37 19 -

Tóm tắt đề cương sinh học phân tử –Trường Đại học Nguyễn Tất Thành
51 26