Nội dung ôn tập sinh học phân tử di truyền | Lí thuyết Sinh học | Trường Đại học khoa học Tự nhiên
Sắc ký protein : cho biết hỗn hợp có bao nhiêu kích thước protein khác nhau, các protein khác nhau có thể có cùng kích thước, khi thực hiện quá trình điện di thì ta sẽ thấy nó ở cùng 1 vạch. (tùy theo nồng độ protein có thể sử dụng sắc ký bản mỏng, sắc ký lỏng cao áp…). Tài liệu giúp bạn tham khảo, ôn tập và đạt kết quả cao. Mời đọc đón xem!
Môn: Sinh học (SH) 16 tài liệu
Trường: Trường Đại học Khoa học tự nhiên, Đại học Quốc gia Hà Nội 1.1 K tài liệu
Tác giả:






Preview text:
ACID NUCLEIC DNA RNA
- Deoxyribonucleic acid- Ribonucleic acid - A T G C- A U G C - Dài- Ngắn
- Nhạy cảm với UV- Ít nhạy cảm với UV hơn
- Bền (cấu trúc sợi đôi xoắn kép)-
Không bền (có ct xoắn kép trong 1 số TH)
- Có trong nhân, ty thể- Có trong nhân, TBC DNA
- Có dạng xoắn kép, 1 đoạn xoắn dài 34 Angstron gồm 10 base.
- Có nhóm phosphate hướng ra bên ngoài ptu DNA
- Cấu trúc : 1 nhóm phosphate, 1 phân tử đường 5C, 1 base
- Liên kết H, liên kết phosphodiester (giữa 2 base trên cùng 1mach)
- NST cấu tạo từ DNA và protein
- Có khả năng tự nhân đôi
- X-ray Diffision : phát hiện cấu trúc tinh thể của DNA bằng cách chiếu tia X-
Gồm các đoạn exon (mang các gen được mã hóa và biểu hiện ra ngoài) và
intron (không được mã hóa và biểu hiện ra ngoài)
- Gen nhảy (mobile) & gen immobile (gen cố định tại 1 vị trí, không thay đổi
vịtrí từ NST này sáng NST khác hoặc những vị trí khác nhau trên cùng 1 NST).
- Coding gen ( đoạn DNA chứa tp quy định cấu
trúcmã hóa của gen, protein) & non-conding (là
trình tự nu không mã hóa ra bất cứ axit amin nào) -
Quá trình điện di ( sự di chuyển của DNA do dòng điện)
+ Cột nhiều vạch là thang đo, 3 cái còn lại là mẫu.
+ DNA di chuyển từ cực âm sang cực dương.
+ Độ sáng của vạch tỉ lệ thuận với DNA
-> Cho thấy kích thước của DNA. Đoạn DNA nào di
chuyển về chiều dương nhanh hơn thì có cấu trúc nhỏ hơn.
- Thí nghiệm Mesels - Stahl : chứng minh DNA tái bản theo nguyên tắc bán bảo toàn. RNA -
Cấu trúc : 1 nhóm phosphate, 1 phân tử đường 5C (nhiều hơn 1O so với
đường của DNA), 1 base (thay T bằng U).
+ Cấu trúc bậc 1 : Sợi RNA đơn
+ Cấu trúc bậc 2 : 1 cấu trúc đôi RNA nằm ở 1 vị trí bất kỳ trên sợi (cấu trúc kẹp tóc và vòng cuốn).
+ Cấu trúc bậc 3 : nút giả & cuống. -
Vai trò : sinh tổng hợp protein & tham gia vào quá trình điều hòa của protein -
Quá trình già hóa RNA : RNA mới được sinh ra phải trải qua quá trình
cắt bỏ intron, nối exon, cố định bằng chuỗi poly-A ở đuôi và cap ở đầu. -
Điện di RNA (tương tự DNA) (do mang điện âm từ gốc phosphate nên
khi đặt vào điện trường RNA & DNA sẽ di chuyển, tùy kích thước sẽ di chuyển
những đoạn ngắn dài ở trên gel) PROTEIN -
Vai trò : chất xúc tác, vận chuyển, cấu tạo và duy trì sự phát triển của cơ
thể (cơ), sự lan truyền tín hiệu (signaling : VD như insullin điều hòa lượng đường trong máu) -
Các loại LK của protein : LK H, LK peptide(giữa các aa), S-S (đặc trưng
cho cấu trúc bậc 3-4), LK ion, tương tác kị nước. -
Các bậc cấu trúc của protein :
+ Bậc 1 : liên kết đơn giản nhất giữa các aa tạo thành chuỗi
+ Bậc 2 : từ chuỗi thông qua liên kết H tạo thành CTB2 là xoắn α và β
+ Bậc 3 : từ bậc 2 thêm LK S-S, LK H, LK ion -> CTB3 (tiểu đơn vị)
+ Bậc 4 : các tiểu đơn vị liên kết với nhau. -
Sắc ký protein : cho biết hỗn hợp có bao nhiêu kích thước protein khác
nhau,các protein khác nhau có thể có cùng kích thước, khi thực hiện quá trình
điện di thì ta sẽ thấy nó ở cùng 1 vạch. (tùy theo nồng độ protein có thể sử dụng
sắc ký bản mỏng, sắc ký lỏng cao áp…) -
Để xác định chính xác loại protein thì ta sử dụng Western Blotting : là
kỹ thuật sử dụng kháng thể để nhận diện protein thông qua hình ảnh. Bước đầu
tiên là tách protein cần nhận diện khỏi hỗn hợp bằng pp điện di (tách theo kích
thước). Bước 2 là chuyển kết quả điện di đó lên 1 màng đặc hiệu. Bước 3 là
đưa màng đặc hiệu đã được chuyển kết quả kết hợp với protein đặc hiệu (sử
dụng kháng thể chuyên nhận diện loại protein đó). -
MALDI - TOF : giải trình tự aa. LIPID
- Chất béo bão hòa và chất béo không bão hòa.
- Được phân thành 8 loại bởi LIPID MAPS
- Chức năng cấu trúc : lớp phospholipid kép trong màng sinh chất. Đầu ưa
nước (gốc phosphate có thể gắn với cholin) đưa ra
ngoài và đầu kỵ nước (các acid béo) quay vào trong, tạo màng linh động.
- Đóng vai trò nhận diện tín hiệu & truyền tín hiệu
- Lipid raft : có cấu trúc linh động trên màng tế
bào,cấu tạo như 1 cụm gồm cholesterol
glycosphingolipids và các protein. Các phần đưa ra bên ngoài sẽ đóng vai trò
là nhận diện tín hiệu và truyền tín hiệu vào bên trong - Có 3 loại sắc ký :
+ Sắc ký phân vùng : là sự phân tách các thành phần giữa 2 pha lỏng (…)
+ Sắc ký trao đổi ion : là quá trình cho phép tách các ion và phân tử phân
cực trên cơ sở ái lực của chúng đối với chất trao đổi ion.
+ Sắc ký hấp phụ : ứng dụng của quá trình hấp phụ lên bề mặt của cột những
protein và loại bỏ những protein không thể hấp phụ lên bề mặt đó, sau đó rửa
giải những protein đã gắn kết trên bề mặt của cột CARBOHYDRATE -
Phân loại : mono, di, oligo, poly (phân nhánh & không phân nhánh) -
Chức năng cấu trúc : thành phần tạo nên nucleotide, thành TB thực vật, nấm,
màng vk. Trên bề mặt TB, glucose liên kết protein tạo nên các cấu trúc đa dạng
về bề mặt TB, những TB cùng loại có thể khác nhau về % glycoprotein để phân
biệt TB này với TB khác) - Quá trình trao đổi chất. -
Truyền tín hiệu : khi kết hợp với glycoprotein -> đóng vai trò như
receptor, nhận diện các phân tử bên ngoài TB -> kích hoạt hệ thống các chất và
lan truyền tín hiệu vào bên trong bằng quá trình phosphoryl hóa các phân tử
bên trong. Tín hiệu sẽ gia tăng dần sau các phản ứng phosphoryl hóa. => tác
động vào bên trong -> tổng hợp các protein mới đáp lại các tín hiệu được nhận
diện bởi glycoprotein trên bề mặt TB. -
Có thể phân loại theo kích thước nhờ phương pháp điện di (phải xử lý
borate trước khi điện di - Các loại sắc ký :
+ Sắc ký bản mỏng : xác định các chất trong TB có độ tan khác nhau trong dung môi..
+ Sắc ký khí : tách và phân tách các chất dễ bay hơi mà không làm phân hủy nó.
+ HPLC : phương pháp sắc ký lỏng phân tích các thành phần tự nhiên nhờ tia tử ngoại. ※
CHƯƠNG 2 : CƠ CHẾ DI TRUYỀN PHÂN TỬ CƠ BẢN
Tính biến động và ổn định của DNA
Tính biến động
Tính ổn định
- Biến tính (2 mạch đơn lk với nhau - Hồi tính (khi nhiệt độ tăng từ 80 đến
bằng lk H, các tác nhân 95℃ làm 2 95℃ DNA bị biến tính, khi hạ nhiệt mạch
tách rời nhau) độ thì lk H hình thành lại, DNA cuộn
xoắn lại như ban đầu) (qt duy trì trạng thái ban đầu
- Đột biến (điểm, đoạn, indel) - Cơ chế sửa sai - Sao chép (lỗi) - Sao chép bán bảo tồn
- Tái tổ hợp - Cấu trúc mạch đôi xoắn kép - Trao đổi chéo - Mobile element
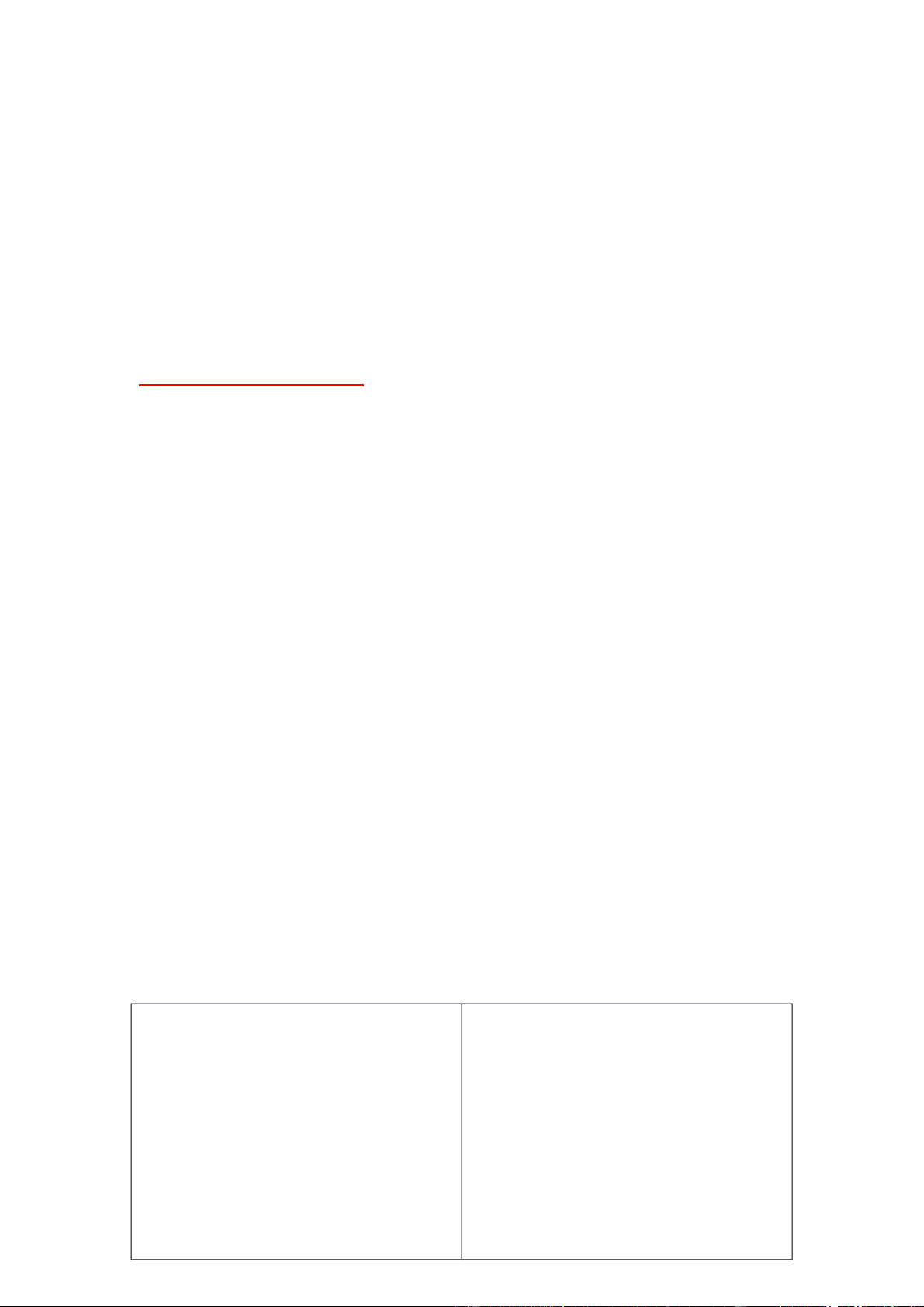
Sự sao chép và sửa sai Sự sao chép :
+ Enzyme khởi động quá trình tháo xoắn : Topoisomerase. + Helicase tháo xoắn + Protein single - strand
binding có vai trò ổn định
cấu trúc mạch đơn, tránh
sợi đơn liên kết lại với nhau ( do DNA có xu
hướng xoắn lại để đạt trạng thái bền vững) + Leading strand tổng
hợp cùng chiều với chiều tháo xoắn của Helicase. + DNA polymerase cần
đầu 3’-OH mới có thể kéo dài được mạch mới. + Lagging strand được
tổng hợp không liên tục. Enzyme primase sẽ tổng
hợp các đoạn mồi là RNA primer, sau đó các đoạn mồi sẽ được kéo dài bởi
DNA polymerase tạo thành các đoạn Okazaki.
+ Sau khi các đoạn RNA primer được DNA polymerase thay thế bằng các
đoạn DNA thì DNA ligase sẽ nối các đoạn Okazaki thành một mạch mới liên tục. DNA Helicase
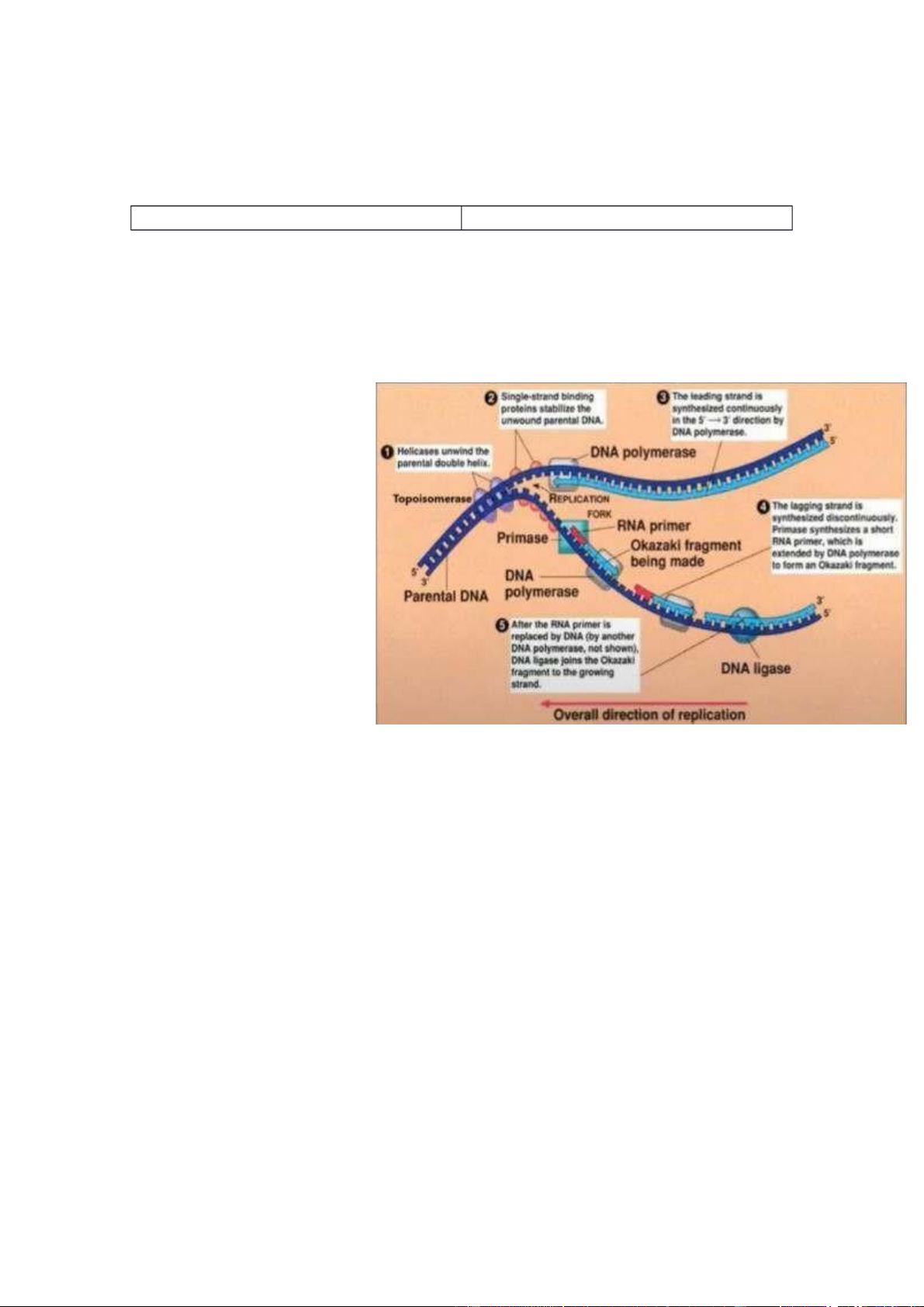
+ Gồm 3 tiểu đơn vị : B, C, D
+ Helicase cắt liên kết H giữa các cặp nucleotide giữa 2 mạch.
+ Hoạt động nhờ năng lượng ATP.
+ 2 tiểu đơn vị RecB và RecD phụ thuộc vào
hoạt động của enzyme ATPase. RecB và
RecD sẽ di chuyển theo 2 chiều trái ngược
nhau ( RecB 3’->5’ ; RecD 5’->3’), trong
quá trình đó nó sẽ cắt liên kết H => tách 2 mạch sợi DNA. DNA Polymerasae
- Khi tổng hợp DNA mới, DNA polymerase
bắt đầu từ đầu 3’ và hướng sự tổng hợp về
phía đầu 5’ bằng cách thêm nucleotide bổ
sung cho khuôn mẫu. DNA polymerase cần 1
nhóm 3’-OH tồn tại trước để bắt đầu tổng
hợp chuỗi và được tạo điều kiện thuận lợi bởi
đoạn DNA hoặc RNA nhỏ gọi là mồi. DNA
polymerase đọc DNA khuôn mẫu và di
chuyển từ đầu 3’ sang đầu 5’ tạo ra sợi DNA
mới theo chiều 5’->3’ .
- Cấu trúc : có 2 cấu trúc. 1 cấu trúc thực hiện
tổng hợp DNA, 1 cấu trúc thực hiện quá trình
chỉnh sửa DNA. Tùy vào chức năng của nó
tại 1 thời điểm mà nó sẽ có cấu trúc tương ứng.
+ Cấu trúc bao mạch khuôn DNA và thực
hiện quá trình thêm nucleotide trong chính cấu trúc đó (chạy trên mạch đơn DNA để tổng hợp).
* Tóm lại : với mạch khuôn, có 1 đoạn mồi gắn ở vị trí đầu 3’, chiều gắn của
mạch mới là 5’->3’ -> có gốc OH ở vị trí kết thúc của nucleotide (vị trí 3’),
nucleotide tự do sẽ gắn đầu 5’ của nó vào 3’ tại vị trí gốc OH. Khi đó, gốc
phosphate ở triphosphate sẽ loại ra gốc là pyrophosphate sinh ra gốc
phosphate tự do. Khi đó, 3 gốc phosphate chỉ còn lại 1 gốc phosphate, gốc
phosphate này sẽ liên kết với nucleotide trước đó ở vị trí 3’, làm cho sợi DNA
được kéo dài theo chiều 5’ đến 3’.
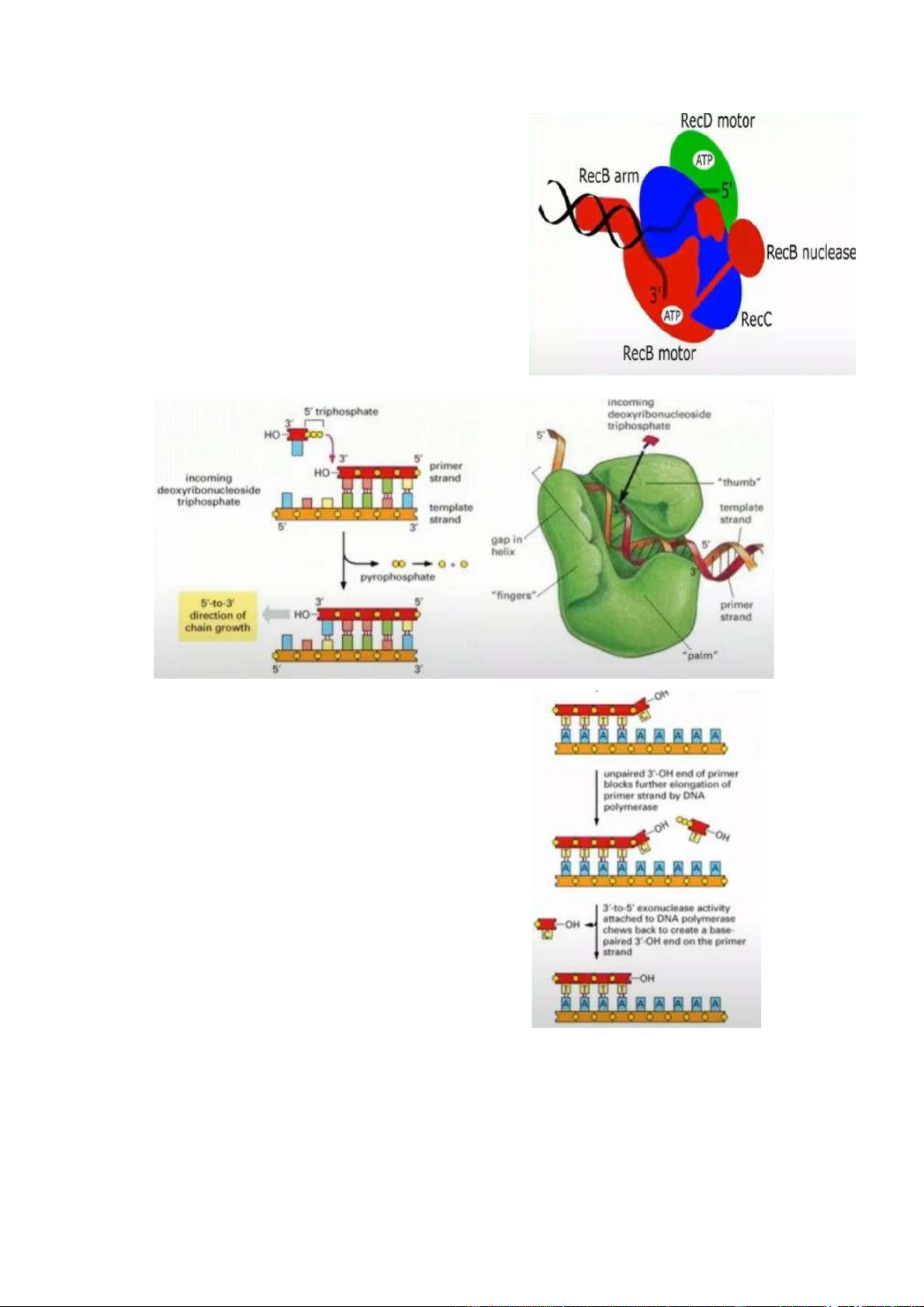
+ Trạng thái chỉnh sửa sai sót (editing) : tùy mỗi loại DNA polymerase mà sẽ
có xác suất sai sót khác nhau. DNA polymerase ở cấu dạng này vẫn trượt trên
mạch khuôn DNA và sẽ nhận dạng được nucleotide vừa gắn là đúng hay sai để chỉnh sửa.
- P : vị trí thực hiện quá trinh tổng hợp, E : vị trí kiểm tra và chỉnh sửa. Nếu
quátrình tổng hợp DNA bằng cách thêm nu vào và nó nhận diện rằng tại vị trí
3’OH nu thêm vào không đúng thì quá trình tổng hợp dừng lại, không cho
phép nucleotide mới được tiếp cận và tổng hợp tiếp tục, ngăn chặn việc bỏ
qua nucleotide bị sai, để sửa
nucleotide đó. Quá trình sửa đó
cũng được thực hiện bởi DNA
polymerase bằng cách loại bỏ
nucleotide bị gắn sai nhờ hoạt động
của exonuclease nằm trong DNA
polymerase. Sau đó nó tiếp tục tổng
hợp ( sợi con đang ở vị trí P, lúc này
DNA polymerase đang gắn các
nucleotide tự do để kéo dài mạch,
sau khi gắn xong nó sẽ đi qua vị trí của ngón cái
để dò lại, nếu có sai sót nó sẽ đưa sợi con xuống vị trí E. Lúc này, exonuclease
sẽ được kích hoạt. Nó sẽ cắt nucleoitide bị sai và thay thế bằng nu đúng. Sau
đó nó sẽ được chuyển trở lại vị trí P và ngón cái sẽ trở lại vị trí như hình
polymerizing). [exonuclease : cắt ngoài mạch vào, endonuclease : cắt giữa mạch] .
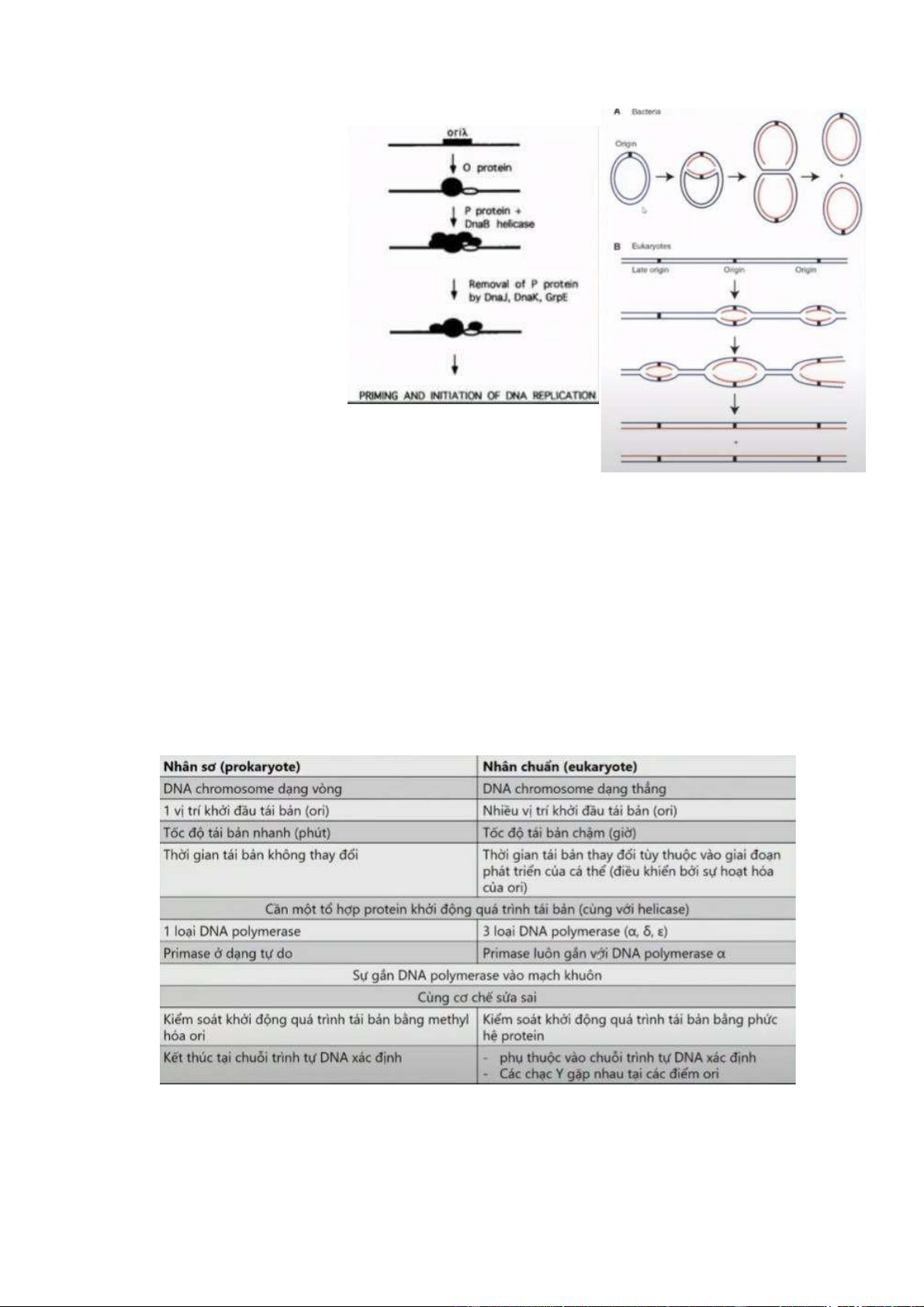
- Trước khi DNA tháo xoắn sẽ có 1 phức hợp để nhận diện vị trí tháo xoắn (
priming and initiation of DNA replication). Nó sẽ nhận diện tại vị trí Ori (vị
trí khởi đầu) . Ở nhân sơ và nhân thực có sự khác biệt về số lượng Ori. DNA
sẽ được khởi động quá trình sao chép bắt đầu từ những Ori.
+ Protein O nhận diện điểm khởi đầu Ori, bắt đầu kêu gọi các protein khác
bao gồm helicase, tham gia tạo thành phức hợp. Sau khi DNAB helicase được
gắn vào, nó sẽ bắt đầu loại bỏ protein P - phức hợp được hình thành tại vị trí
Ori. Khi phức hợp gắn tại vị trí Ori, helicase sẽ thực hiện chức năng tháo
xoắn. (Ở sinh vật nhân sơ chỉ có 1 điểm Ori. Từ điểm Ori đó nó sẽ nhận diện
bởi phức hợp khởi động quá trình sao chép. Tại điểm Ori sơi DNA được tách
ra và tại vị trí cắt được gọi là chạc chữ Y. Chạc chữ Y
sẽ được mở rộng dần và khi nó đi hết một vòng của sợi DNA mạch khuôn - đồng
thời tổng hợp nên sợi DNA con - nó tìm lại
vị trí Ori ban đầu thì 2 DNA con được tổng hợp theo nguyên tắc
bán bảo tồn sẽ được tách ra. Ở TB nhân
thực thì có nhiều điểm Ori, trong đó có những điểm Ori không hoạt động hoặc cách rất xa các điểm
Ori khác -> Có mô hình hoạt động giống domino :
điểm Ori thứ 1 được nhận diện và kích hoạt thì nó sẽ
làm cho điểm Ori kế tiếp nó được kích hoạt. Toàn bộ
những điểm Ori này sẽ được kích hoạt và thực hiện
quá trình tháo xoắn DNA. Khi tổng hợp mạch con
theo nguyên tắc bán bảo tồn thì những điểm này gặp
nhau và nó sẽ kết thúc quá trình tổng hợp DNA con.
Mô hình thứ 2 : từng điểm Ori được kích hoạt, không
cần phản ứng dây chuyền, nhưng đích cuối cùng vẫn
là sự gặp gỡ của các chạc chữ Y và các điểm Ori ->
quá trình sao chép dừng lại ).
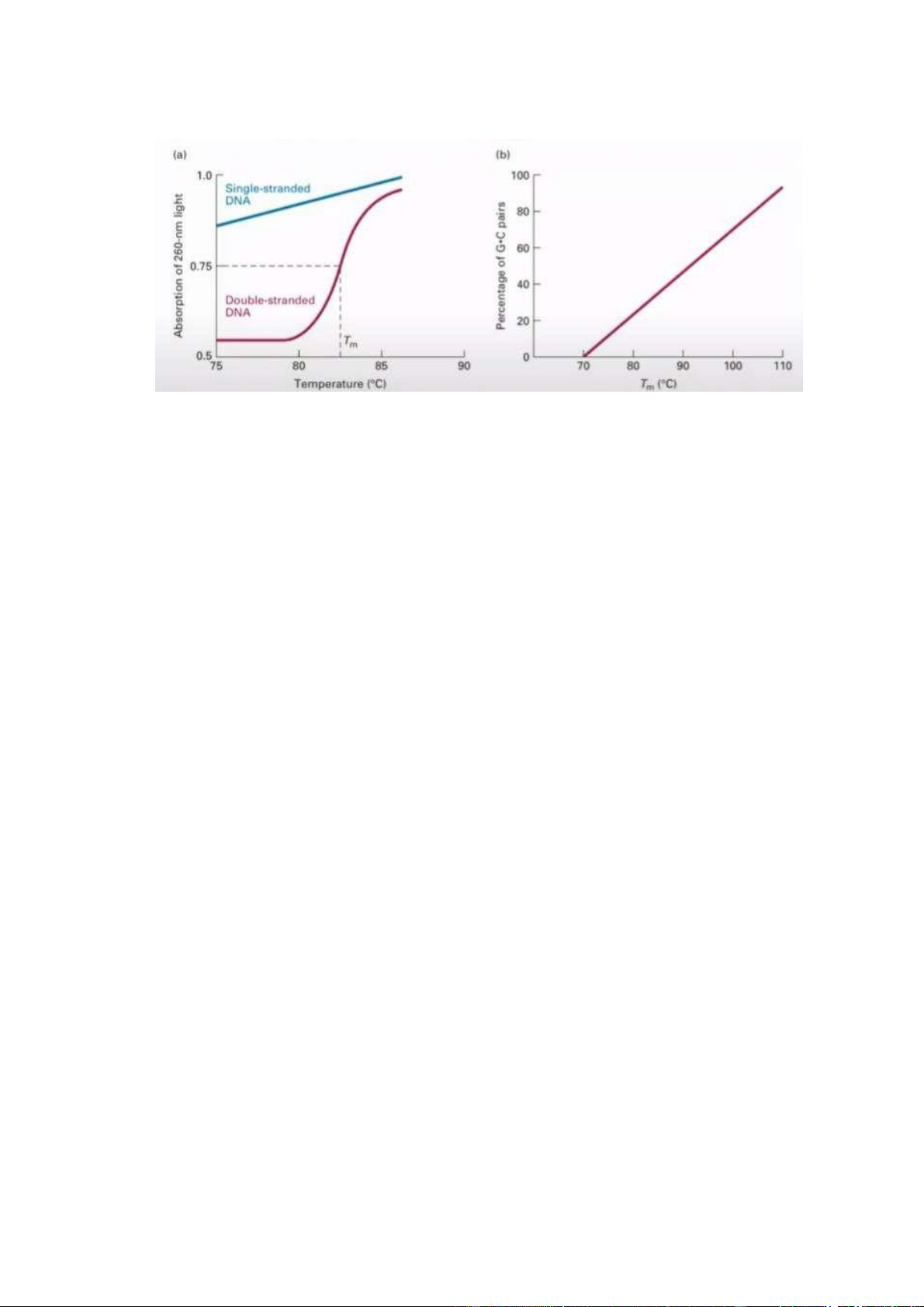
Biến tính và hồi tính
- Tại vị trí nhiệt độ nóng chảy, DNA chuyển từ trạng thái mạch kép sang trạng
thái mạch đơn. Nếu nhiệt độ lớn hơn nhiệt độ nóng chảy, DNA sẽ bị biến tính
làm cho cấu trúc mạch đôi thành cấu trúc mạch đơn (biến tính). Khi hạ nhiệt
độ xuống dưới nhiệt độ nóng chảy, DNA sẽ trở lại trạng thái mạch đôi (hồi
tính). Tính biến tính là khác nhau giữa DNA, phụ thuộc nhiệt độ nóng chảy
(nhiệt độ nóng chảy phụ thuộc vào phần trăm G-C - % G-C càng cao thì nó
càng bền với nhiệt độ, nhiệt độ nóng chảy của DNA đó càng lớn).
- Độ hấp thụ bước sóng ở 260nm. Trong phòng TN có thể ứng dụng bằng việc
đo nồng độ DNA hoăc acid nucleic thông qua sử dụng máy đo UD (??) (đo độ
hấp thụ ánh sáng ở bước sóng 260nm) để ước lượng nồng độ DNA. Các
biến động của acid nucleic
- Các nhân tố ảnh hưởng đến tính biến động của acid nucleic :
+ Nhân tố bên ngoài : tia bức xạ (tia gamma, tia UV), chất hóa học ( có khả
năng oxi hóa mạnh) => ảnh hưởng trực tiếp lên cấu trúc DNA khiến
nuclotide bị biến đổi (đột biết điểm), nếu tác nhân quá mạnh có thể trực tiếp
làm đứt gãy sợi DNA và quá trình sửa chữa không thể hoàn thành -> mất luôn
sợi DNA hoặc có thể sửa chữa được nhưng không chính xác (bằng cách so
sánh trình tự nucleotide mà nó đang có với DNA tương đồng - quá trình đó
làm nucleotide không còn chính xác như ban đầu)
+ Tác nhân bên ngoài nhưng gián tiếp gây ra tín hiệu bên trong TB làm acid
nucleic bị biến động. VD : bản thân các hạt nano không thể làm thay đổi cấu
trúc DNA. Nhưng khi TB nằm trong môi trường có chứa các hạt nano đó, hạt
nano đi vào bên trong thì nó làm cho bên trong tế bào đó sinh ra rất nhiều chất
oxy hóa. Các chất oxy hóa này sẽ tác động ngay chính TB và DNA làm đứt
gãy hoặc đột biến DNA.
+ Một số biến động acid nucleic là kết quả của quá trình sửa sai. VD : quá
trình sửa sai loại bỏ các base, đáng lẽ đã hoàn tất nhưng vì 1 số lý do quá trình
này không được thực hiện 1 cách hoàn chỉnh -> làm tồn tại các nucleotide bị
sai -> trình tự acid nucleic bị đảo lộn.
+ Sự tồn tại của gene nhảy : làm thay đổi thứ tự acid nucleic tại vị trí gene nhảy chèn vào.
+ Quá trình phân chia TB có hiện tượng trao đổi chéo và tái tổ hợp -> tạo giao tử khác biệt.
+ Trong quá trình tái bản có sử dụng polymerase - hoạt động nhưng vẫn có xác suất bị sai. Base excision repair
- Nucleotide nhận diện bị sai nhờ vào
DNA glycosylase và tương ứng với
từng loại nucleotide thì sẽ có từng loại
DNA glycosylase tương ứng để nhận
diện. Và khi nhận diện nó sẽ loại bỏ
base của nucleotide đó. Nucleotide đó
chỉ còn gốc phosphate và phân tử
đường. Khi này, AP endonuclease sẽ
cắt bỏ liên kết phosphodiester với
nucleotide kế tiếp. Tiếp đó, AP lyase
sẽ loại bỏ hoàn toàn 2 thành phần còn
lại của nu đã bị mất base (giai đoạn
loại bỏ hoàn toàn nu bị sai) -> xuất
hiện vị trí bị trống. Lúc này, DNA
polymerase và ligase sẽ lấp đầy vị trí
bị trống và hoàn thành các liên kết với
mạch còn lại => DNA được sửa chữa.
Double strand break repair
+ Tái tổ hợp tương đồng : sợi DNA màu xanh da trời bị đứt gãy, 1 trình tự
DNA tương đồng (không giống hoàn toàn đoạn bị đứt gãy, có khả năng bắt
cặp với nhau) không bị đứt gãy màu xanh lá cây -> nó sẽ lấy đoạn có khả
năng bắt cặp đó và sao chép lại đoạn bị đứt gãy dựa trên khuôn là trình tự
mà nó bắt cặp. Kết thúc quá trình là sợi DNA bị đứt gãy đó sẽ chứa trình tự
của DNA tương đồng với DNA đã bị đứt gãy.
+ Quá trình nối đoạn đơn : sợi DNA có đoạn đầu màu cam, đoạn sau màu
vàng, khúc giữa bị cắt. Các đoạn màu cam, vàng chứa trình tự có thể bổ sung
được cho nhau. Khi bị cắt đứt thì màu vàng có thể bắt cặp với màu cam. Khi
bắt cặp như vậy thì đoạn màu cam ở trên và đoạn màu vàng ở dưới bị dư ra -
> phần dư ra sẽ bị loại bỏ -> kết quả : 1 đoạn trình tự mới chứa cả màu cam và màu vàng.
+ Nối không tương đồng : Quá trình này cần sự hiện diên của enzyme.
Enzyme sẽ bám vào 2 đầu bị đứt và nối trực tiếp lại với nhau mà không cần sự
hiện diện của đoạn tương đồng.
- 1,2 ít gây tổn thương hơn so với 3. Nhưng xác xuất để trở lại trình tự ban đầu
là rất thấp. So về độ tổn thương thì 3 gây ra tổn thương cho tế bào và trình tự acid nucleic nhiều nhất.
Sửa chữa chạc chữ Y sụp đổ
- Chạc chữ Y là nơi helicase bám vào vàthực
hiện quá trình tháo xoắn. Ở đây có 2 mạch
là mạch nhanh (tổng hợp theo chiều mở
của helicase) và mạch chậm (tổng hợp
ngược chiều mở helicase).
- [Điểm màu đen là điểm đột biến]. Nếu
điểm đột biến nằm gần vị trí tháo xoắn,
chạc chữ Y có thể không tiến đi nữa mà sẽ
dừng lại -> dừng lại quá trình sao chép -> chạc chữ Y sụp đổ.
+ TH1 : sửa chữa để làm mất điểm đột
biến. Chuỗi tổng hợp nhanh gặp điểm đột
biến -> lùi điểm đột biến từ phần nhánh
vào phần chân chạc chữ Y. Khi dó, đoạn mạch chậm (không bị ảnh hưởng bởi
điểm đột biến) đã tổng hợp được 1 đoạn dài hơn đoạn bị đột biến -> trở thành
cái khuôn cho mạch bị ảnh hưởng bởi điểm đột biến. Trong quá trình chạc chữ
Y được lùi lại, cả 2 mạch đều được tổng hợp với cấu trúc tương đồng. Lúc
này, chạc chữ Y bắt đầu tiến tới và đi qua điểm đột biến -> trong mạch con
không còn chứa điểm đột biến.
+ TH2 : lờ đi. Mạch con không sửa mà đi qua luôn -> giữ lại điểm đột biến.
+ TH3 : sửa chữa bằng phương pháp lấy trình tự đoạn bên dưới. Nó sẽ lấy
mạch bên dưới không bị đột biến trao đổi với mạch bên trên -> tạo ra cấu trúc
mới : 1 DNA con chứa trình tự mạch khuôn mẹ và 1 DNA mẹ chứa trình tự của mạch DNA con. Yếu tố di động
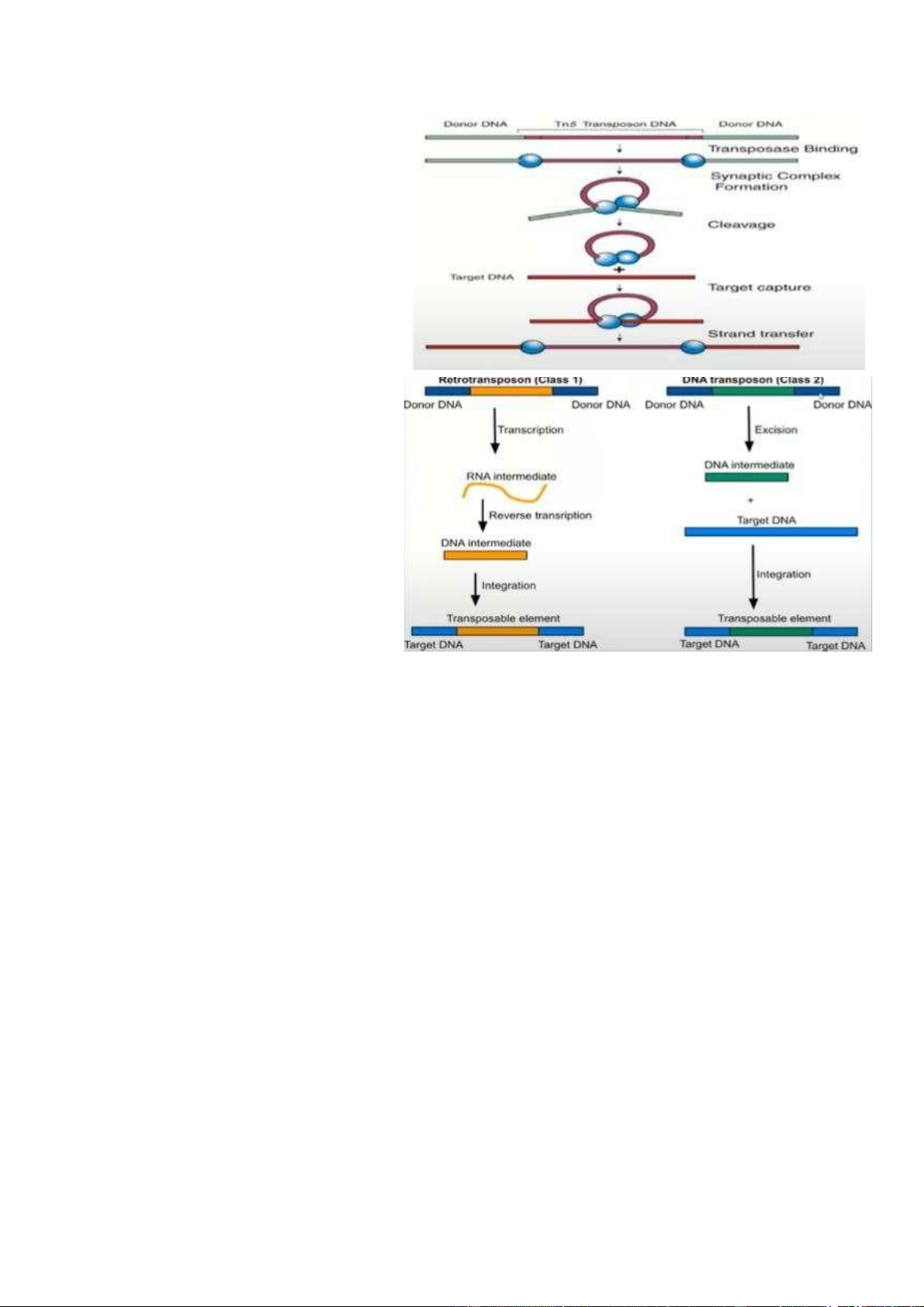
- DNA Transposon bản chất là DNA. - Khi nó thực hiện quá
trìnhchèn, nó sẽ chèn trực
tiếp vào DNA đích. Việc
chèn trực tiếp đó sẽ thông
qua việc nhận diện, cắt và
dán của enzyme transposase. - Gen nhảy có thúc đẩy
quátrình tiến hóa hay không
còn phụ thuộc vào tác động
của quá trình chèn đó vào
gen đích. Nếu quá trình chèn
đó không gây chết cá thể thì
nó mới là nguyên liệu đóng
góp cho quá trình chọn lọc và tiến hóa.
- Phân loại thứ hai của gennhảy là retrotransposon
(nhân tố chuyển vị ngược). - Bản chất của
retrotransposonlà RNA, thực hiện quá trình sao chép ngược.
- Bản chất retrotransposon làphải có quá trình phiên mã ngược từ RNA sang
DNA, nhưng khi nó nằm ở gen đích thì nó phải ở dạng DNA. Donor DNA
(DNA cho) là trình tự DNA đã được sao chép ngược từ retrotransposon (dưới
dạng RNA). Donor DNA phải thực hiện quá trình phiên mã để tạo RNA trung
gian -> RNA trung gian phiên mã ngược thành DNA trung gian -> DNA trung
gian mới chèn vào DNA nhận => Nguồn của gen nhảy không bị mất đi.
* Giống nhau : đều mang yếu tố di động, đều được hoạt động nhờ enzyme cắt và dán. * Khác nhau : …
- Một số yếu tố di động khác : Ở vi khuẩn có mang gen kháng sinh. Qua quá
trình giao nạp và tái tổ hợp, nó sẽ chuyển plasmid từ vi khuẩn cho sang vi
khuẩn nhận. Hoạt động của yếu tố di động là plasmid không phụ thuộc vào
gen nằm trong genome, trong khi những cái được đề cập ở trên thì tương tác
vào chính genome của cá thể.
- Yếu tố di động có khả năng di động khắp genome và sẽ chèn 1 cách ngẫu
nhiên vào bộ gen. Việc ngẫu nhiên đó vừa có những điểm thuận lợi (nguyên
liệu cho quá trình tiến hóa) vừa có những điểm bất lợi (thông thường gây ra
phản ứng tiêu cực, đa phần sự chèn của gen nhảy vào genome sẽ gây chết cá
thể). Nếu cá thể không bị chết thì sự chèn của gen nhảy sẽ tồn tại trong quần
thể, và trong điều kiện không có môi trường chọn lọc thì nó sẽ tiếp tục có xu
hướng tích lũy những biến đổi đó. Đến 1 lúc nào đó đạt đến giá trị cực đại thì
nó sẽ làm cá thể bị chết.
- Khi cơ thể gặp nguy cơ lớn với điều kiện bất lợi, nó sẽ tìm cách kiểm soát và
cân bằng bất lợi đó để đảm bảo sao cho cá thể được hoạt động 1 cách tốt nhất
=> Tồn tại những cơ chế chống lại hoạt động của gen nhảy.
- Một số cơ chế chống yếu tố di động (mobile elements) ở động vật có vú :
+ KRAB/KAP1 sẽ làm biến đổi histone bằng cách methyl hóa DNA, làm biến
đổi vị trí nhận diện các yếu tố di động.
+ Hoạt động của nhóm protein APOBECs : loại bó gốc amin -> phá hủy cấu
trúc của yếu tố di động.
+ Nhóm PIWI protein, piRNA : làm phân cắt hoặc methyl hóa yếu tố di động.
+ Yếu tố di động càng tiến hóa để có khả năng liên tục chèn vào hệ thống
genome -> genome cũng tiến hóa thông quá việc lựa chọn protein APOBECs,
KRAB/KAP1 để chống lại sự chèn của yếu tố di động. ※
CHƯƠNG 3 : SỰ PHIÊN MÃ VÀ ĐIỀU HÒA PHIÊN MÃ
Cấu trúc gene
- Gồm 3 phần : Promoter (vùng khởi động cho quá trình phiên mã được diễn ra
- TTGACA…17nu-TATAAT), RNA coding sequence (vùng mã hóa cho
RNA), Terminator (vùng kết thúc)
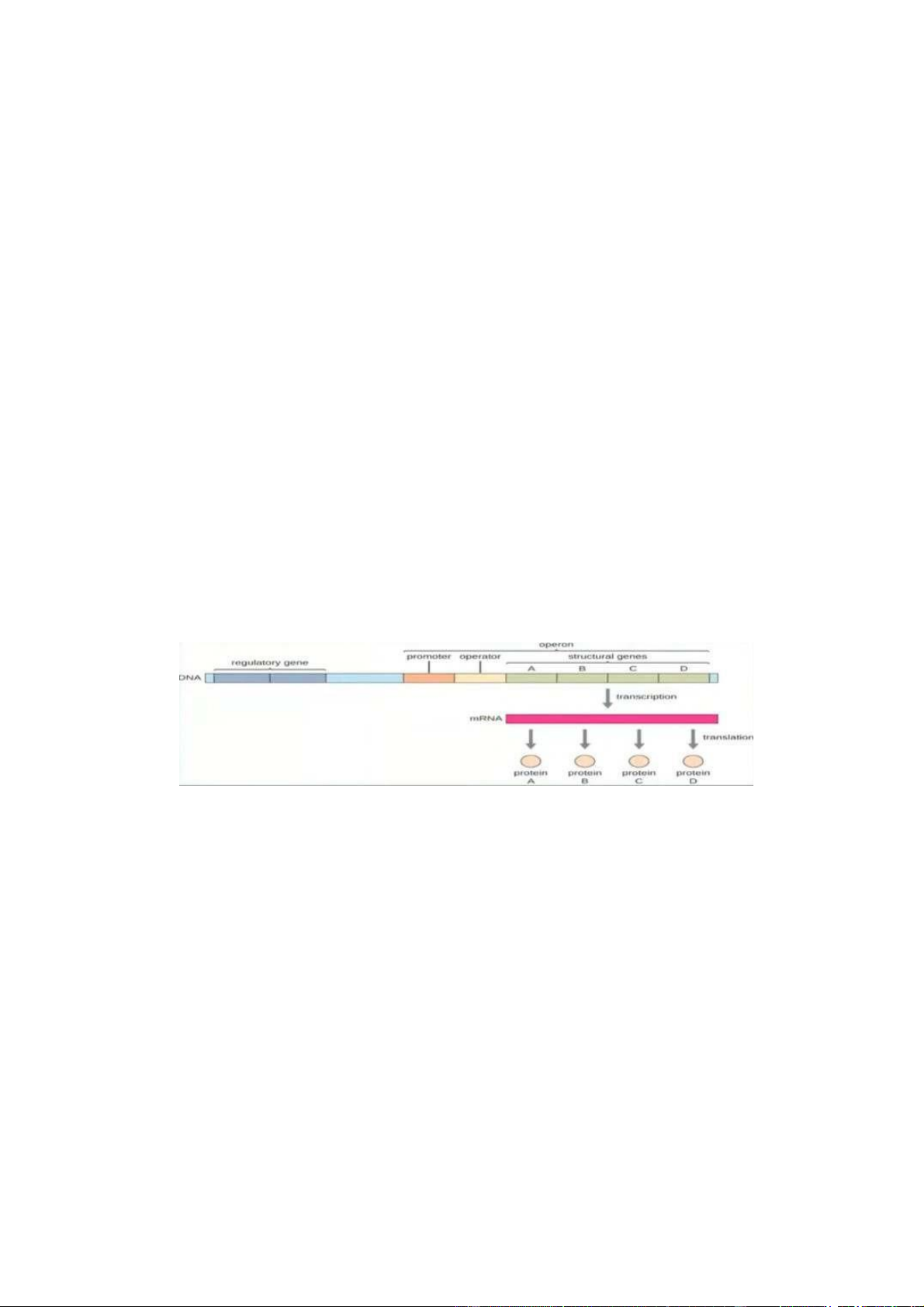
- Cấu trúc gene sinh vật nhân sơ : +
Các gen tồn tại dưới dạng cụm Operon. VD : trên hình operon chứa 4 gen cấu
trúc A,B,C,D. 4 gen cấu trúc này được điều khiển và vận hành bởi 2 thành phần
trước nó là promoter và operator. Ngoài ra hoạt động của promoter và operator
còn chịu ảnh hưởng của 1 số gen điều hòa. Các gen A,B,C,D được đặt liền kề
nhau mà không bị phân cách bởi các đoạn intron như ở sinh vật nhân chuẩn.
Khi promoter và operator hoạt động thì cả 4 gen cấu trúc đều được tổng hợp.
Tương tác giữa RNA polymerase và DNA-Promoter
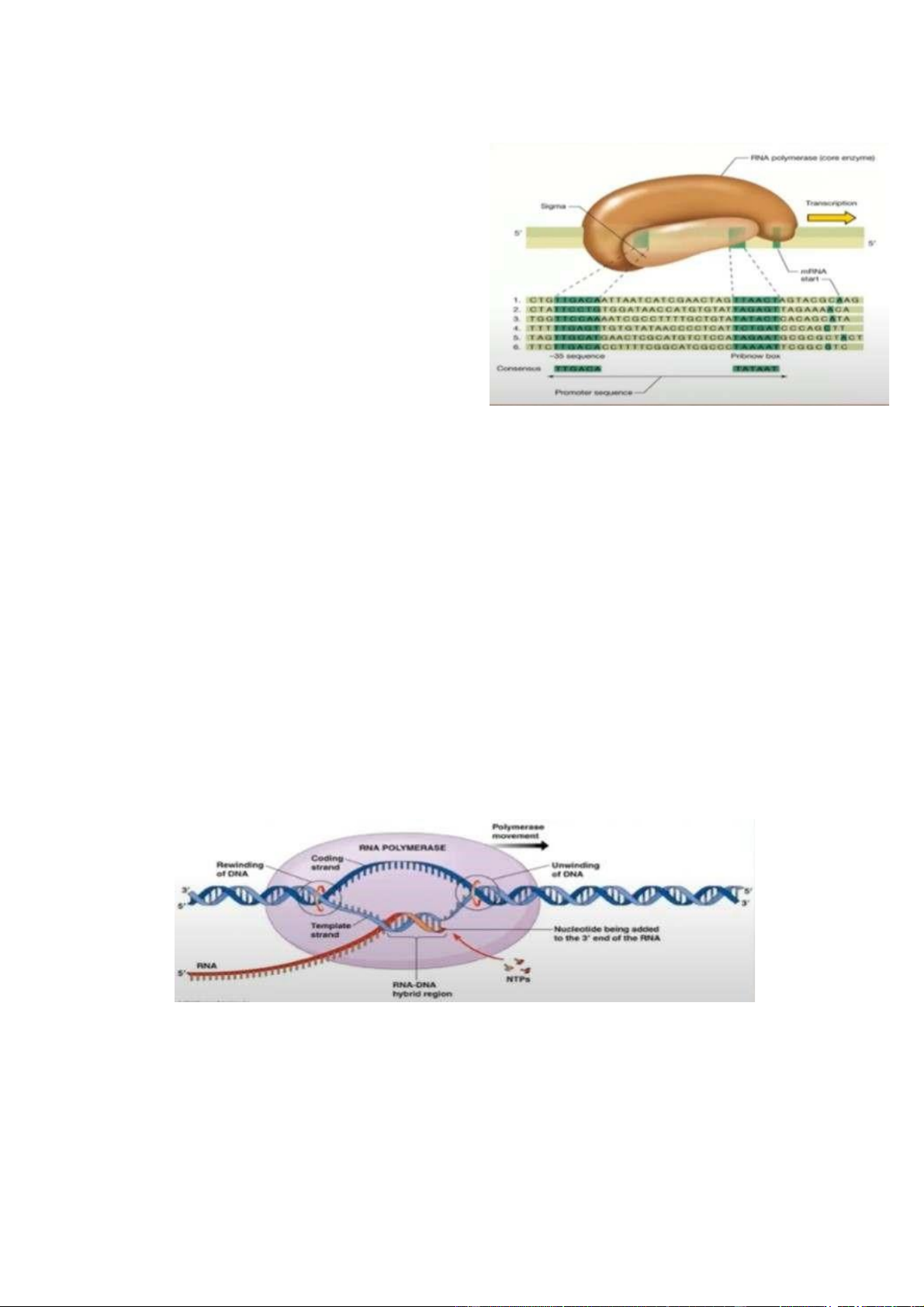
- Để gen thực hiện quá trình phiên mã,
RNA polymerase phải tương tác, gắn kết
với vùng khởi động (promoter). - Quá
trình nhận diện và bám dính của RNA
polymerase và promoter được đảm bảo
bởi trình tự bảo thủ.
+ Factor σ trên RNA polymerase gắn vào vùng promoter.
+ RNA polymerase (core enzyme) gắn vào factor σ.
+ Holoenzyme thực hiện tháo xoắn sợi DNA.
+ RNA polymerase bắt đầu từ vị trí -10 (vị trí 1 là vị trí nu đầu tiên của đoạn
mã hóa nên RNA, từ vị trí đó trở về trước - thuộc vùng promoter - sẽ được
đánh dấu âm) để xác định vị trí phiên mã.
+ Factor σ rời khỏi phức hợp, enzyme bắt đầu phiên mã. Kéo dài
- Sau giai đoạn nhận diện vị trí khởi động của quá trình phiên mã tại vị trí
promoter thì RNA polymerase sẽ thực hiện kéo dài để tổng hợp ra sợi
nucleotide quy định trình tự của RNA -> quá trình kéo dài.
+ RNA polymerase sẽ di chuyển trên sợi đơn DNA (là kết quả của quá trình
tháo xoắn của holoenzyme). Khi RNA polymerase di chuyển trên sợi đơn DNA
nó sẽ tổng hợp đồng thời RNA.
+ Quá trình tổng hợp này được thực hiện trong 1 cấu trúc gọi là bong bóng
phiên mã. Bong bóng phiên mã này chạy tới đâu thì nó sẽ giải phóng bớt RNA
đã được tổng hợp trước đó. Quá trình này diễn ra liên tục cho đến khi nó gặp tín hiệu kết thúc. Kết thúc
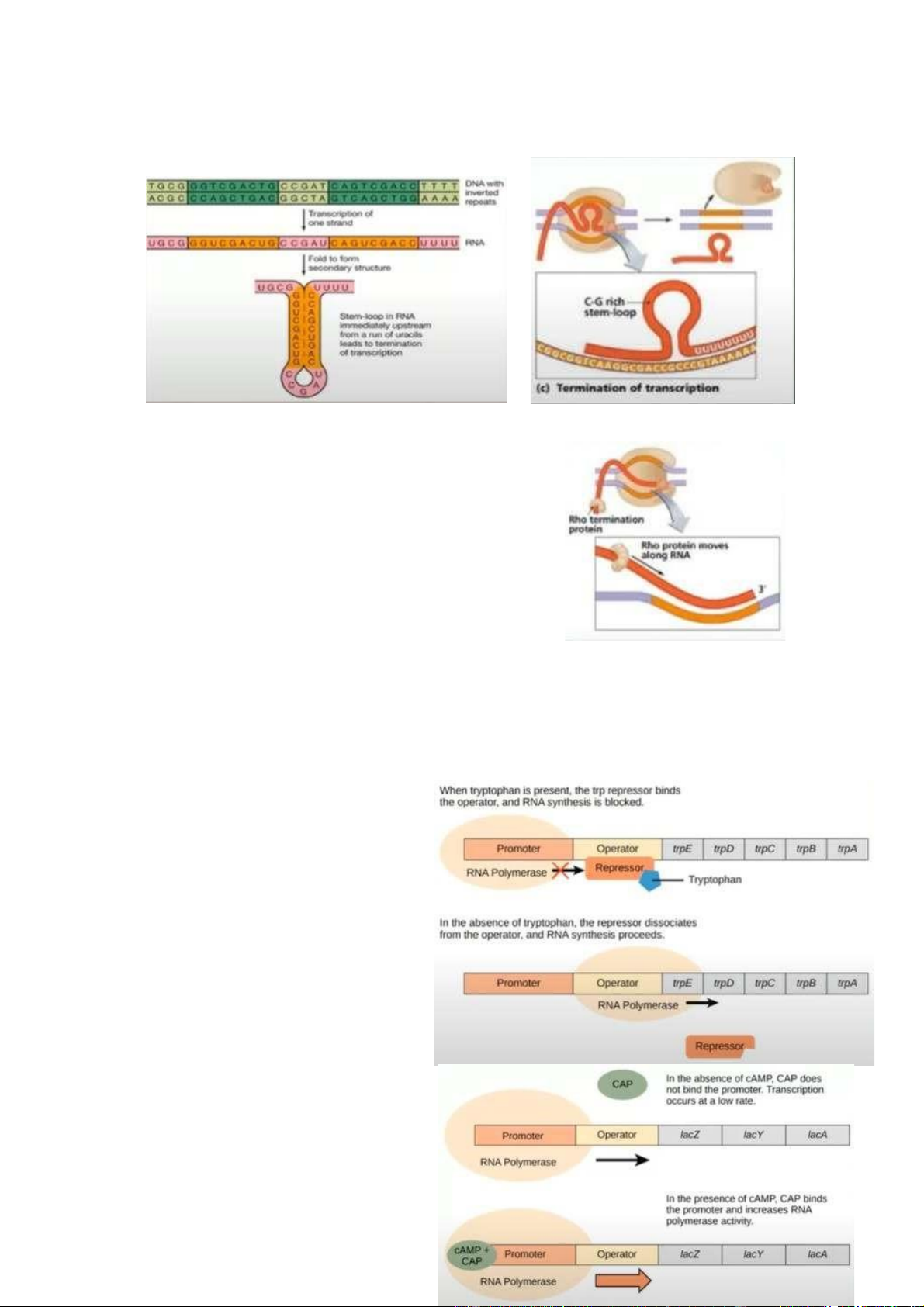
- Loại 1 : không phụ thuộc protein Rho
+ Tín hiệu kết thúc của DNA có cấu trúc bổ sung để tạo 1 cấu trúc như cấu trúc
kẹp tóc (stem-loop) có rất nhiều liên kết G-C - là liên kết khá bền. Bong bóng
phiên mã chạy dọc theo cấu trúc DNA và khi nó gặp cấu trúc kẹp tóc (trình tự
DNA phiên mã ra trình tự RNA, trong bong bóng phiên mã RNA liên kết với
nhau tạo thành cấu trúc kẹp tóc) đưa ra tín hiệu để phức hợp RNA polymerase
rời khỏi DNA, giải phóng hoàn toàn RNA đã được tổng hợp.
- Loại 2 : phụ thuộc protein Rho
+ Protein Rho sẽ gắn vào vùng chứa nhiều C và ít
G trước vị trí kết thúc.
+ Protein Rho đóng vai trò giống helicase (tháo
xoắn) - nó sẽ tháo liên kết giữa RNA và DNA, và
nó sẽ tách rời RNA với DNA.
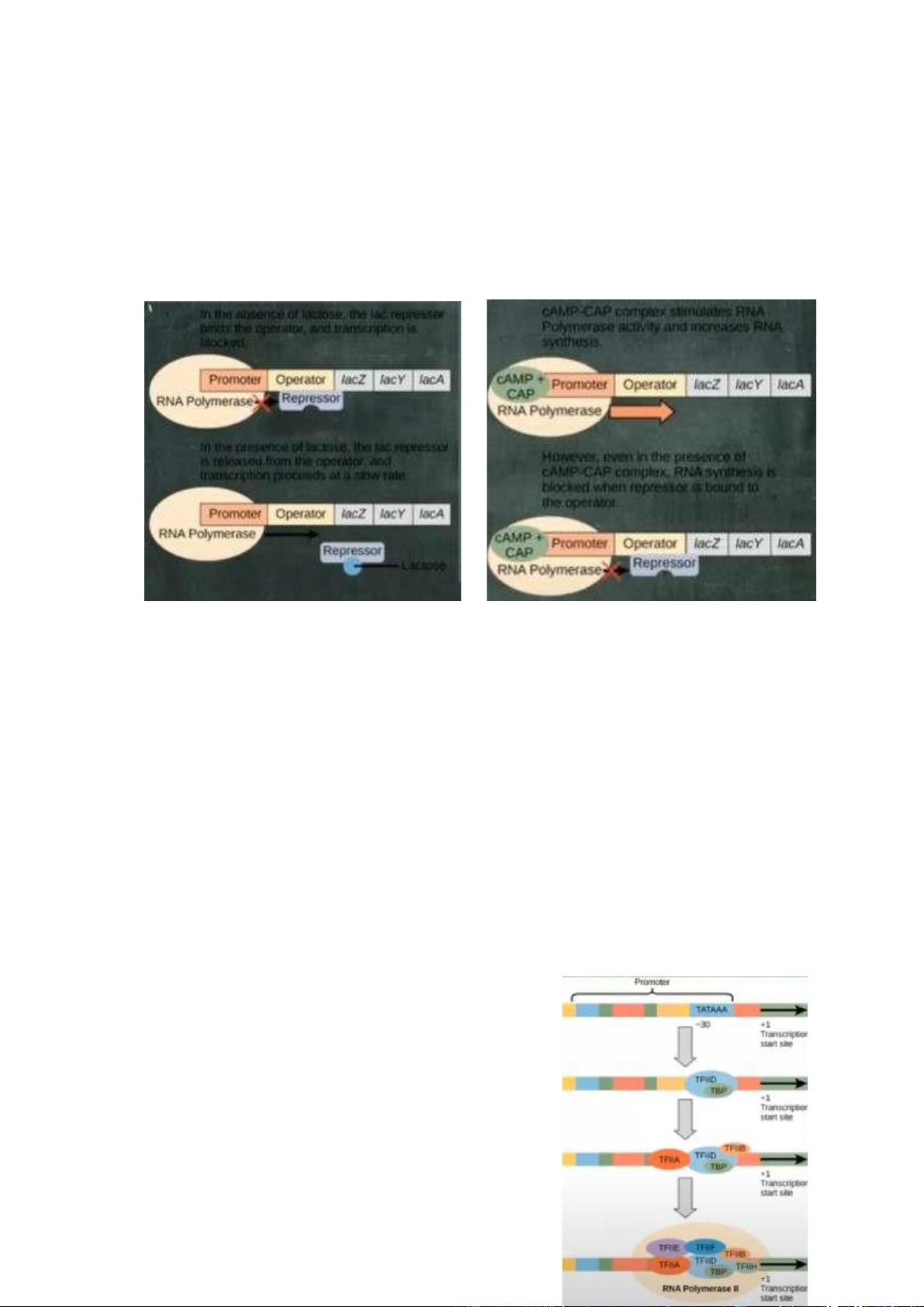
Điều hóa phiên mã ở sinh vật nhân sơ Có 3 cơ chế : -
Ức chế : được đảm bảo bởi nhân tố repressor, gắn vào vùng vận hành
(operator). ngăn cản phiên mã gen đích đối với tín hiệu bên ngoài. (Tryptophan operon)
+ Operon này điều hòa quá trình
tổng hợp tryptophan nên yếu tố
điều hòa là Trp. Khi có sự hiện
diên của Trp tức tế bào không cần
tổng hợp Trp nữa -> ức chế quá
trình tổng hợp Trp : là quá trình
gắn kết giữa Trp với repressor gắn
vào operator - là sự cản vật lý
khiến RNA polymerase không thể
trượt trên sợi DNA, khiến cho các
gen quy định tổng hợp Trp bị dừng lại. -
Kích hoạt : được đảm bảo
bởi activator, gắn vào vùng khởi
động (promoter), gia tăng phiên mã gen đích đối với tín hiệu bên ngoài. (CAP -
Catabolite Activator Protein operon)
+ Khi có sự hiện diện của CAP thì quá trình tổng hợp gen đích được nhiều
hơn. Ở điều kiện không có CAP thì quá trình tổng hợp vẫn diễn ra nhưng với mức độ rất thấp. -
Cảm ứng : được đảm bảo vởi inducer, ngăn cản/gia tăng phiên mã gen
đích tùy vào nhu cầu của tế bào và sự hiện diện của cơ chất. (Lac operon)
+ Chất cảm ứng ở đây là Lac. Nếu không có Lac thì repressor sẽ gắn vào vùng
operator. Nếu có Lac thì Lac sẽ gắn vào repressor và lôi kéo nó ra khỏi operator
và cụm gen tổng hợp Lac ở đằng sau sẽ được thực hiện, tuy nhiên sẽ thực hiện
1 cách yếu ớt. Khi nó nhận thêm tín hiệu CAP thì quá trình này diễn ra mạnh
mẽ -> lượng lactose trong môi trường giảm đi -> yếu tố cảm ứng không có khả
năng gắn kết vào repressor -> repressor quay lại gắn vào operator -> quá trình
tổng hợp nên Lac đằng sau operator bị dừng lại
=> Sự hiện diện của chất cảm ứng quan trọng hơn sự hiện diện của CAP.
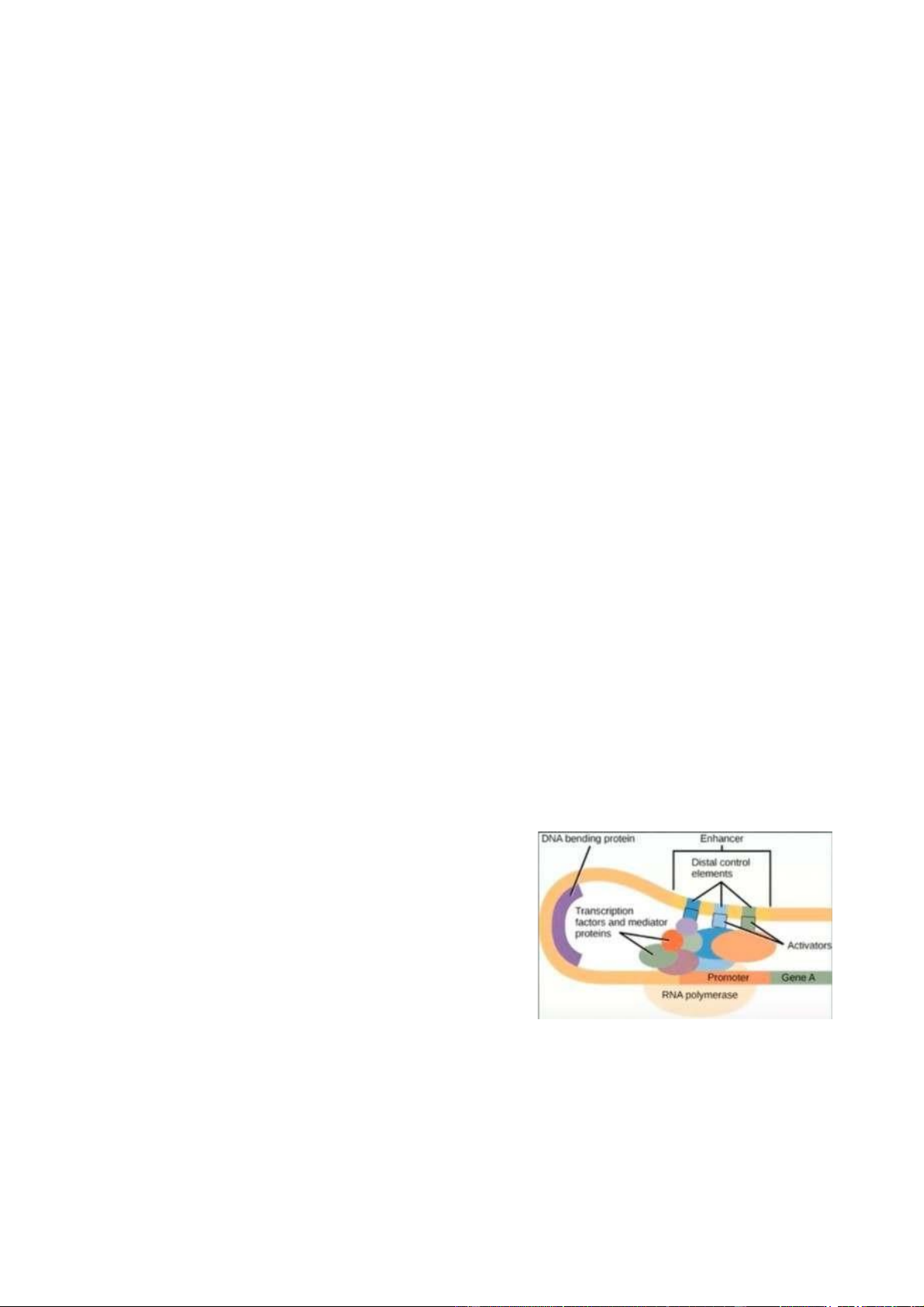
Quá trình phiên mã sinh vật nhân chuẩn - Khởi động -
3 loại RNA polymerase tham gia vào quá
trìnhkhởi động : I ( phiên mã rRNA, trừ 5S
rRNA), II (phiên mã pre-mRNA, microRNA,
long-coding RNA - là những RNA điều hòa quá
trình phiên mã dịch mã), III (phiên mã 5S pre-rRNA, pre-tRNA, small nuclear pre-RNAs) -
Yếu tố phiên mã (transcription factor) gắn vào vùng khởi động
(promoter), gọi polymerase đến gắn kết vào phức hợp tiền khởi động PIC (Pre- initiation complex). -
Nó sẽ được thực hiện tại vị trí TATA box gắn với TBP (TATA - binding
protein), kêu gọi các TFIID/A/B/E/F/H và RNA polymerase để thực hiện quá
trình phiên mã ở mức cơ bản. -
Khi có sự hiện diện của phức hợp này, quá tình phiên mã sẽ được gia
tăng hoặc giảm thiểu do sự tương tác tiếp tục với activator và repressor. Kéo dài -
Đầu tiên phải loại bỏ cấu trúc nucleosome -> phơi ra đoạn DNA. Khi đó
RNApolymerase sẽ tiến hành tổng hợp theo các bước như là tháo xoắn, thêm
ribonu tự do theo chiều 5’->3’. Bong bóng phiên mã hình thành liên kết giữa
RNA và DNA khuôn, độ dài của vùng này chỉ 8 nu nên khi tổng hợp nu mới nó
sẽ phải bật các nu cũ ra để duy trì chiều dài của mạch. -
Phiên mã ở sinh vật nhân chuẩn cần sự hiện diện của 1 protein để loại bỏ
protein histone ra khỏi nucleosome. Kết thúc -
RNA polymerase I : trên DNA khuôn có trình tự đặc biệt nhận biết yếu
tố TTF-1 ( Transcription Termination Factor for RNA polymerase I) -> thông
báo cho polymerase biết để kết thúc quá trình phiên mã tại đó. -
RNA polymerase II : trên DNA cũng chứa trình tự vùng cắt ( cleavage
site) giới hạn bởi AAUAAA - 40 đến 60 nu - vùng giàu GU. Đồng thời nó
cũng gắn thêm đuôi polyA trên đầu 3’ của RNA để tăng tuổi thọ, giúp nó ổn
định hơn - RNA polymerase III : cơ chế chưa được xác định. Điều hòa phiên mã
- Enhancer : gia tăng phiên mã
- Repressor : giảm, ngăn cản phiên mã- Các
yếu tố này có khả năng nằm xa - cách rất xa
so với gen mà chúng tác động. Để nó có khả
năng điều hòa cụm gen đích thì DNA bắt
buộc phải thay đổi cấu trúc 3D của nó để
enhancer hoặc repressor có khả năng tương tác với RNA polymerase.
Điều hòa hậu phiên mã -
Già hóa RNA vừa được tổng hợp : loại bỏ intron - những đoạn phân cắt
RNA mã hóa protein thành những đoạn tách rời nhau - nhờ vào các phức hợp
ribonucleoprotein nhận biết và cắt intron, nối exon lại với nhau. -
Đi kèm với già hóa RNA là ổn định cấu trúc RNA : ổn định bằng cách
thêm vào đầu 5’ cấu trúc cap GTP hoăc thêm vào đuôi 3’ trình tự polyA. Ngoài
ra cấu trúc UTR (Untranslated region) có thể nằm ở đầu 5’ hoặc 3’ - là nơi gắn
kết với RBP ( RNA-binding protein). Tùy vào RBP mà làm tăng giảm độ bền của RNA. -
Cơ chế điều hòa hậu phiên mã quy định khả năng tồn tại hoặc quy định
khả năng tổng hợp protein trong quá trình dịch mã thông qua biến đổi cấu trúc
RNA : micro-RNA (những RNA có kích thước rất nhỏ, khoảng từ 20~200 nu),
thường tồn tại dưới dạng cấu trúc kẹp tóc được tổng hợp ở trong nhân. Sau đó
được vận chuyển ra ngoài nhân và thực hiện quá trình già hóa, tạo ra 1 cấu trúc
cho phép gắn kết bằng liên kết bổ sung trên sợi RNA đích. + Có 2 dạng gắn kết :
1) Nếu như quá trình gắn kết trong phức hợp RISC này gắn kết và tồn tại ở
trên cấu trúc mRNA -> làm cồng kềnh cấu trúc 3D của mRNA -> ngăn chặn
quá trình di chuyển vật lý của ribosome trên sợi mRNA, hạn chế lượng protein được tổng hợp ra.
2) Cũng cấu trúc RISC với micro-RNA gắn trên sợi mRNA và cơ chế miễn
dịch của chính tế bào đó nhận diện đây là cấu trúc RNA kép, dự đoán là cấu
trúc của virus -> phân hủy hoàn toàn mRNA đó -> giảm mRNA kéo theo lượng
protein được tổng hợp bị giảm sút.
CÒN PHÂN CỦA 9 NHÓM THUYẾT TRÌNH
Tài liệu liên quan:
-

ÔN TẬP ĐỘNG VẬT HỌC ĐỘNG VẬT CÓ XƯƠNG SỐNG - BIO HK201
29 15 -

Xét nghiệm phân mảnh DNA tinh trùng và hỗ trợ trong sinh sản | Lý thuyết sinh học | Trường Đại học khoa học Tự nhiên
179 90 -

Tổng hợp các câu trắc nghiệm hay về môn sinh học cần nhớ | Trắc nghiệm sinh học | Trường Đại học khoa học Tự nhiên
406 203 -

Tổng hợp tóm tắt lý thuyết sinh học hay nhất | Lý thuyết sinh học | Trường Đại học khoa học Tự nhiên
416 208 -

Tổng hợp những công thức sinh học hay cần nhớ | Lý thuyết sinh học | Trường Đại học khoa học Tự nhiên
419 210