Carbohydrates Metabolism Part 1 - Glycolysis, TCA Cycle
Tài liệu học tập môn Biochemistry (BT005) tại Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh. Tài liệu gồm 79 trang giúp bạn ôn tập hiệu quả và đạt điểm cao! Mời bạn đọc đón xem!
Môn: Biochemistry (BT005) 10 tài liệu
Trường: Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh 2 K tài liệu
Tác giả:














































Preview text:
lOMoARcPSD|364 906 32 Carbohydrates Metabolism
Glycolysis Citric Acid Cycle Electron Transport Chain Gluconeogenesis 1 1 lOMoARcPSD|364 906 32 Sugars: Glucose & the Carbohydrates 2
Sugars provide us with instant energy, feed our brains, direct
proteins to their destinations, and communicate the identity of
our cells. But they also make some people ill when they eat ice
cream and can poison our cells when present in large quantities. What are sugars?
• Saccharides – carbohydrates
• General chemical formula – CH2O
• Simple – monosaccharides • Disaccharides
• # carbons = # H2O • 2 monosaccharides combined
• glucose, fructose, mannose,
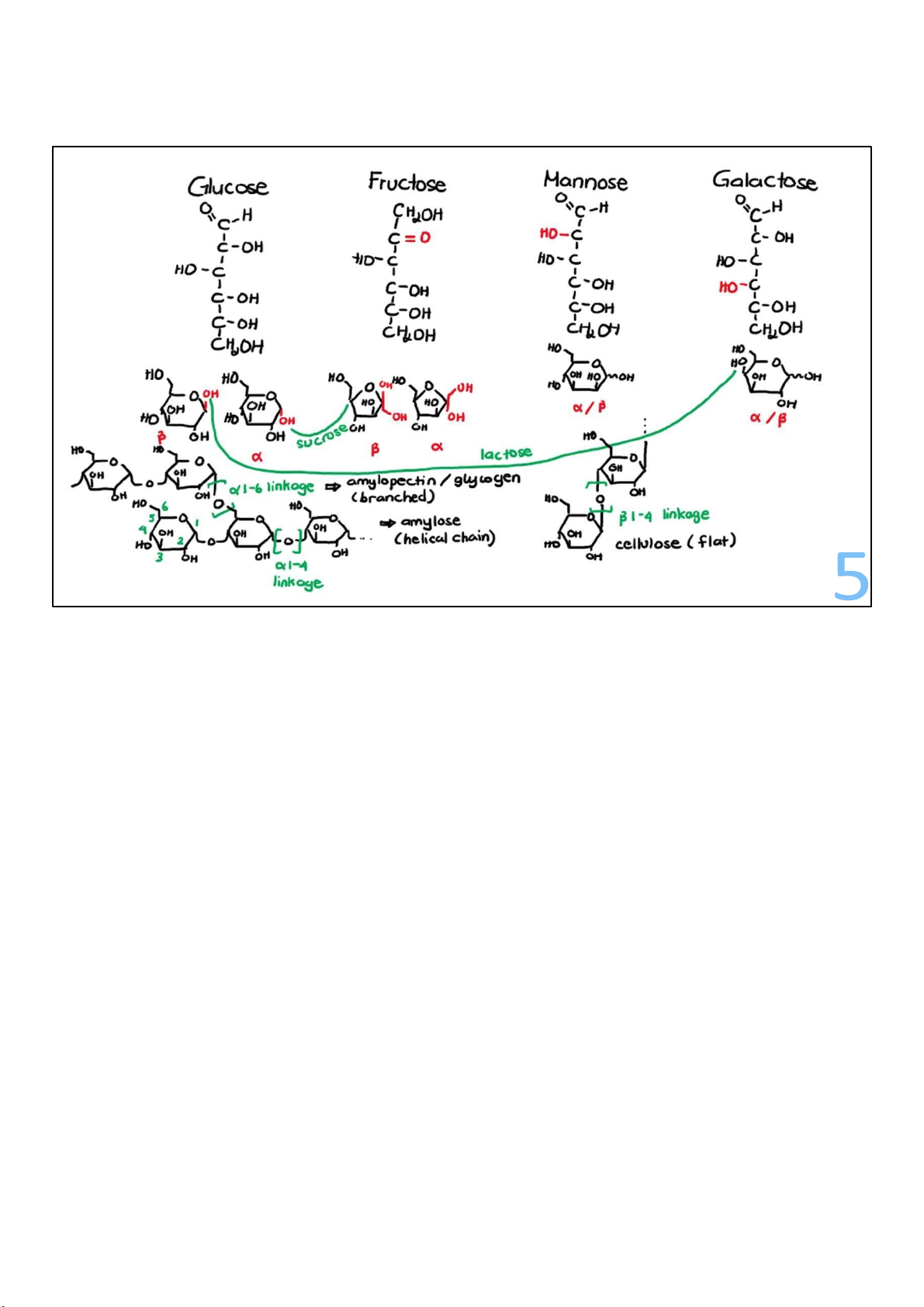
• sucrose (table sugar) = glucose galactose – (CH2O)6 or + fructose C6H12O6 2 lOMoARcPSD|364 906 32 3
Biochemically, sugars are saccharides, a group of molecules
with a particular chemical makeup—a simple carbohydrate.
The general chemical formula for sugars is CH2O, with one
carbon atom for every H2O water molecule.
In simple sugars, or monosaccharides, the number of carbons
in the molecule equals the number of H2Os. The common
monosaccharides we eat—glucose, fructose, mannose, and
galactose—each have 6 carbons and 6 H2Os, giving them each
the formula C6H12O6. They only differ in how they’re organized.
Combine any 2 monosaccharide sugars and you get a
disaccharide. Glucose combined with fructose is ordinary table
sugar, sucrose. Of all the sugars traveling in and around your 3 lOMoARcPSD|364 906 32
cells, fructose is the second most abundant sugar, after
glucose, and the 2 can be readily interconverted. lOMoARcPSD|364 906 32 3 Carbohydrates are polymers
• Poly = many; mer = parts polymers = large molecules from many simpler units
• Simple sugar – building blocks complex carbs
• 3 – 10 simple sugars: oligosaccharides
• > 10 simple sugars: polysaccharides • Energy storage • Structural support
• glycogen (animals) • cellulose (plants)
• starch (plants) • pectin (plants) 4
Simple sugars are building blocks for complex carbohydrates. If
a few simple sugars, between 3 and 10, get linked together,
they are called oligosaccharides.
Longer polymers, typically with more than 10 simple sugars,
are called polysaccharides, which have properties that make
them the ideal energy storage for life, and also work well as structural molecules.
In animals, the sugar polymer is a very large polysaccharide
called glycogen. Plants make a mix of polysaccharides we refer
to as starch (amylose & amylopectin). lOMoARcPSD|364 906 32
Plants don’t make glycogen, but they do use sugars to make
other big carbs beside starch. Very similar to starch, but with a 4
crucial difference, is a cousin polymer called cellulose,
and there are also pectins, which are polymers of modified sugars.
All these molecules differ in the ways the sugars are
organized in the polymer, or the bonds between them. 4 lOMoARcPSD|364 906 32 5 lOMoARcPSD|364 906 32 6 lOMoARcPSD|364 906 32 Why do sugars appeal to us?
• Sweet receptors in tongue (and guts too!) linked to pleasure centre in brain • Sugar = rewards Energy
• Sugar full of energy (not the most energy-dense molecule though)
Fat – twice as much energy, but water-insoluble
special package take time to use (not quick)
• Sugar great for immediate energy 7
We have sweet receptors on our tongues, which are linked to
pleasure centers in our brains. We also have sweet receptors
in our guts. Our bodies reward us for eating sugar and
enthusiastically encourage us to eat more. But why?
Sugars are full of energy, and eating high-energy foods is a simple recipe for survival.
Sugars are not the most energy-dense molecules in the body.
That distinction goes to fat, which, gram for gram, stores
more than twice as much energy as sugar. But fats are
waterinsoluble, so they must be packaged in complexes to
travel in the bloodstream. Then, on arrival at their cellular 7 lOMoARcPSD|364 906 32
destinations, fats have to be separated from their carrier
molecules before they can be used. 8 lOMoARcPSD|364 906 32
This all takes time, so fats are a poor choice for quick energy.
When we need immediate energy, sugars are the go-to
source. Not only are sugars a source of ready energy,
but their water solubility means they can easily travel in the bloodstream.
Virtually every cell in every organism uses sugar in a
central metabolic pathway—a part of metabolism that’s
so essential we can’t live without it.
This is why sugar is bundled together to make glycogen
in our liver and muscle cells.
But at any given time, we only have about a 24-hour
supply of sugar present in our bodies, stored as
glycogen. If you did not eat for a day or so, you’d use up
all your glycogen. And that can be problematic,
especially because our brains strongly prefer glucose to
fuel activities. If blood glucose levels fall too low, the
resulting hypoglycemia can be dangerous—even fatal. 7 lOMoARcPSD|364 906 32
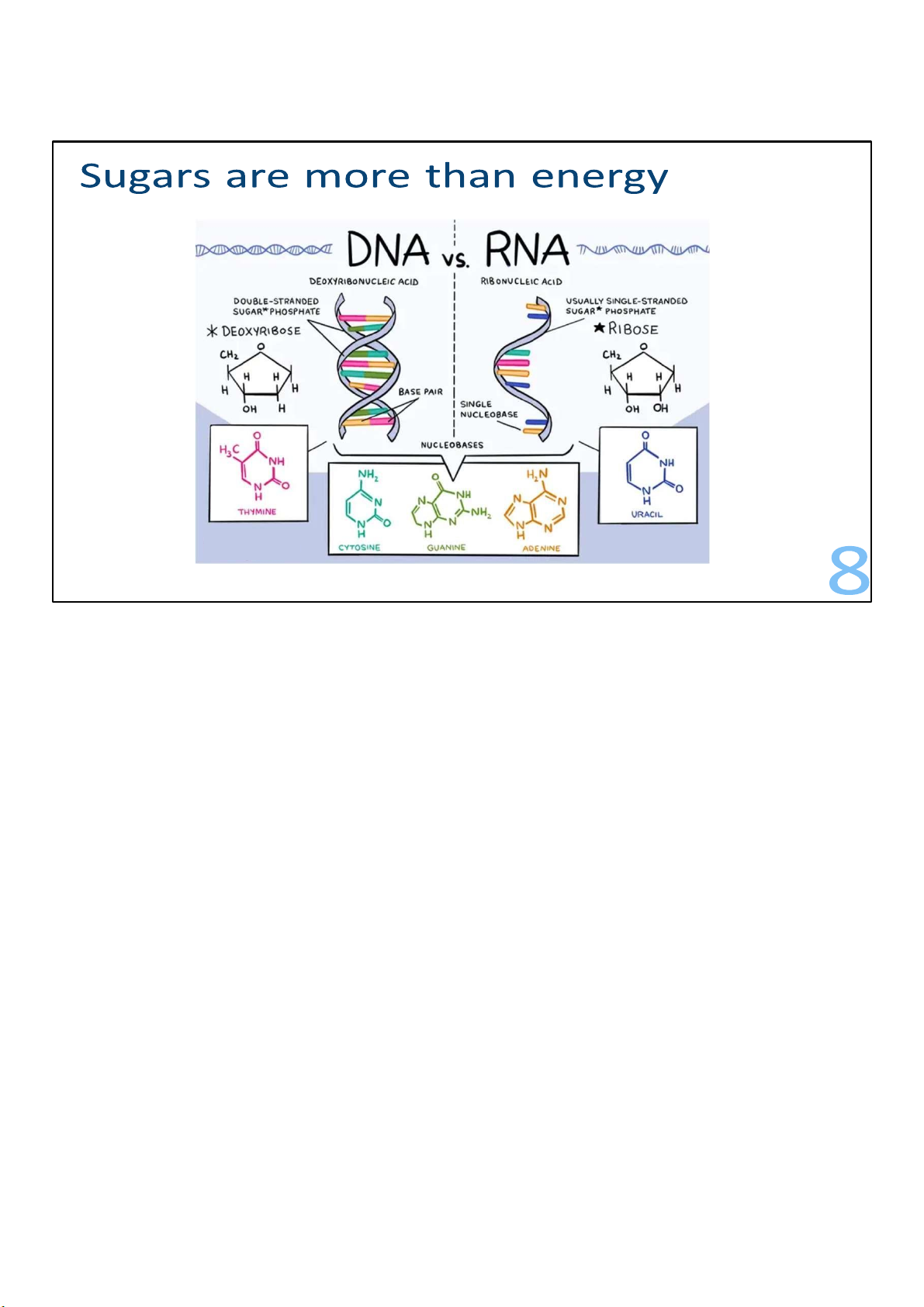
But sugars are used for more than energy. For example, ribose
contributes to the structure of RNA, and a related sugar called
deoxyribose contributes to the structure of DNA. 8 lOMoARcPSD|364 906 32
Proteins in animal cell membranes have small groups of
sugars, called glycans, attached covalently to them in such a
way that the sugars stick out on the outside of the cell. These
are oligosaccharides that are often branched, so they look like
little trees growing out of the membrane proteins. These are examples of glycoprotein.
The kinds of oligosaccharide trees attached to the membrane
proteins serve as a sort of cellular bar code, identifying what
type of cell it is. Proteins with oligosaccharides attached to
them are called glycoproteins, and each cell membrane has thousands of them. 9 lOMoARcPSD|364 906 32
One example of glycoproteins that are involved in cellular ID
are the blood group antigens or markers, which help determine 10 lOMoARcPSD|364 906 32
blood types. Whichever kind of blood group antigens are
found on your cell membranes determines whether you
are A, B, or AB. If your blood group is O, your cells don’t have any of the markers. 9 lOMoARcPSD|364 906 32 Glycolysis 10
Let’s go back to the story of energy with glycolysis.
Metabolic pathways are essentially identical for humans and
blue whales—or any other organism. Consequently, if you
learn a metabolic pathway for one organism, you’ve already
learned a pathway that’s in almost every other organism. A metabolic pathway
• 2 stages, 10 steps (in cytoplasm) • From glucose (6C) • To 2 pyruvate (3C) oxidation
release of energy (stored in phosphate bonds of ATP) 11 10 lOMoARcPSD|364 906 32
Cells act on glucose in 10 steps in glycolysis. Glucose contains 6
carbons and gets broken down to 2 identical molecules of
pyruvate, each with 3 carbons.
Other sugars can be oxidized here as well. In the process, glucose is oxidized.
This involves a release of energy, which is partly stored in the phosphate bonds of ATP.
The glycolysis pathway starts in the cytoplasm of cells. Stage 1: Energy investment Steps 1-5 12
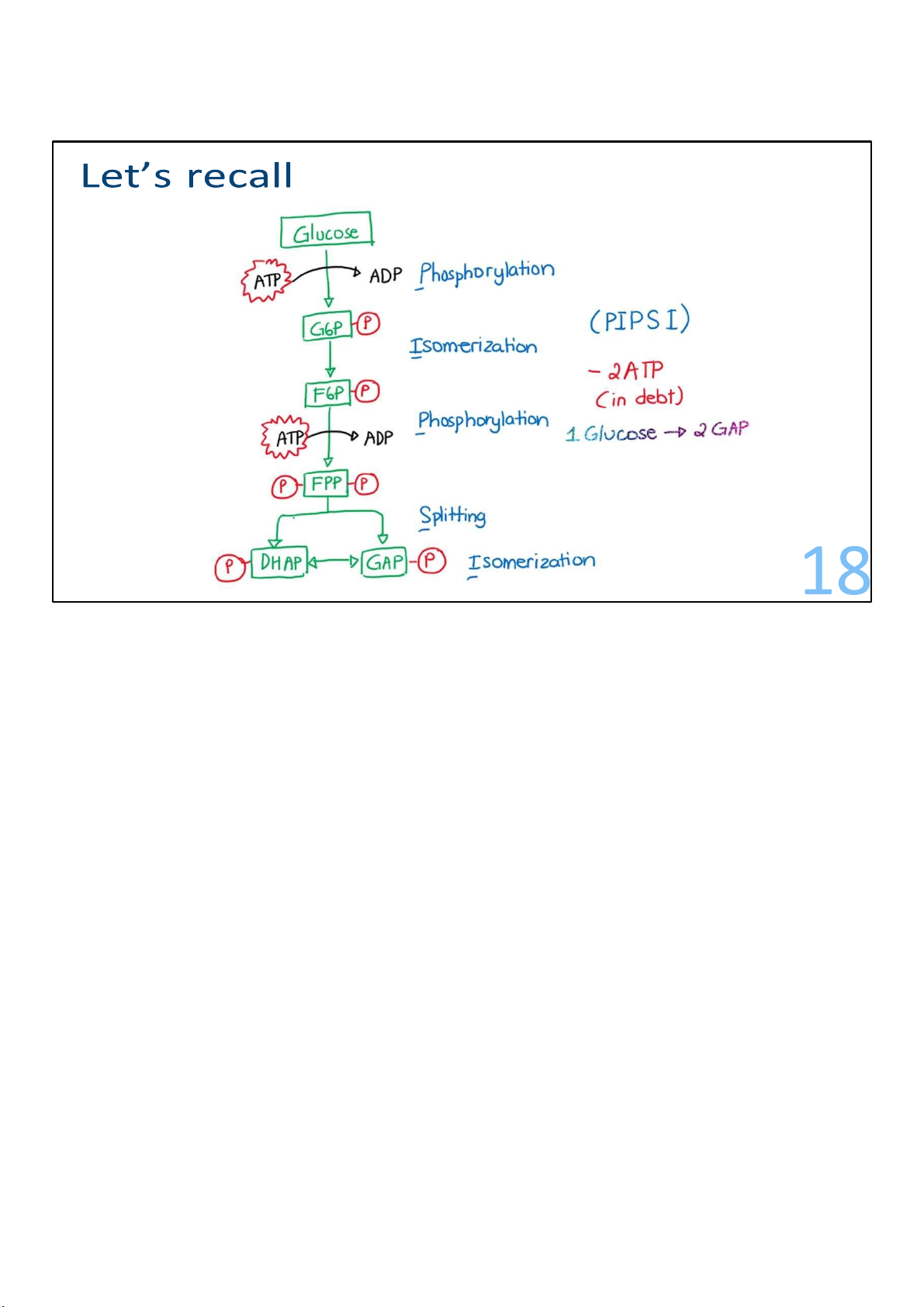
From glucose to 2 glyceraldehyde-3-phosphate
In this early stage, the 6C glucose is phosphorylated and cleaved to yield two molecules of the
triose glyceraldehyde-3-phosphate. This process actually consumes ATP. We actually lose ATPs
during this stage, but don’t worry, we’ll get them right back. 11 lOMoARcPSD|364 906 32
We’ll take a look at each step, and then sum them up at the end.
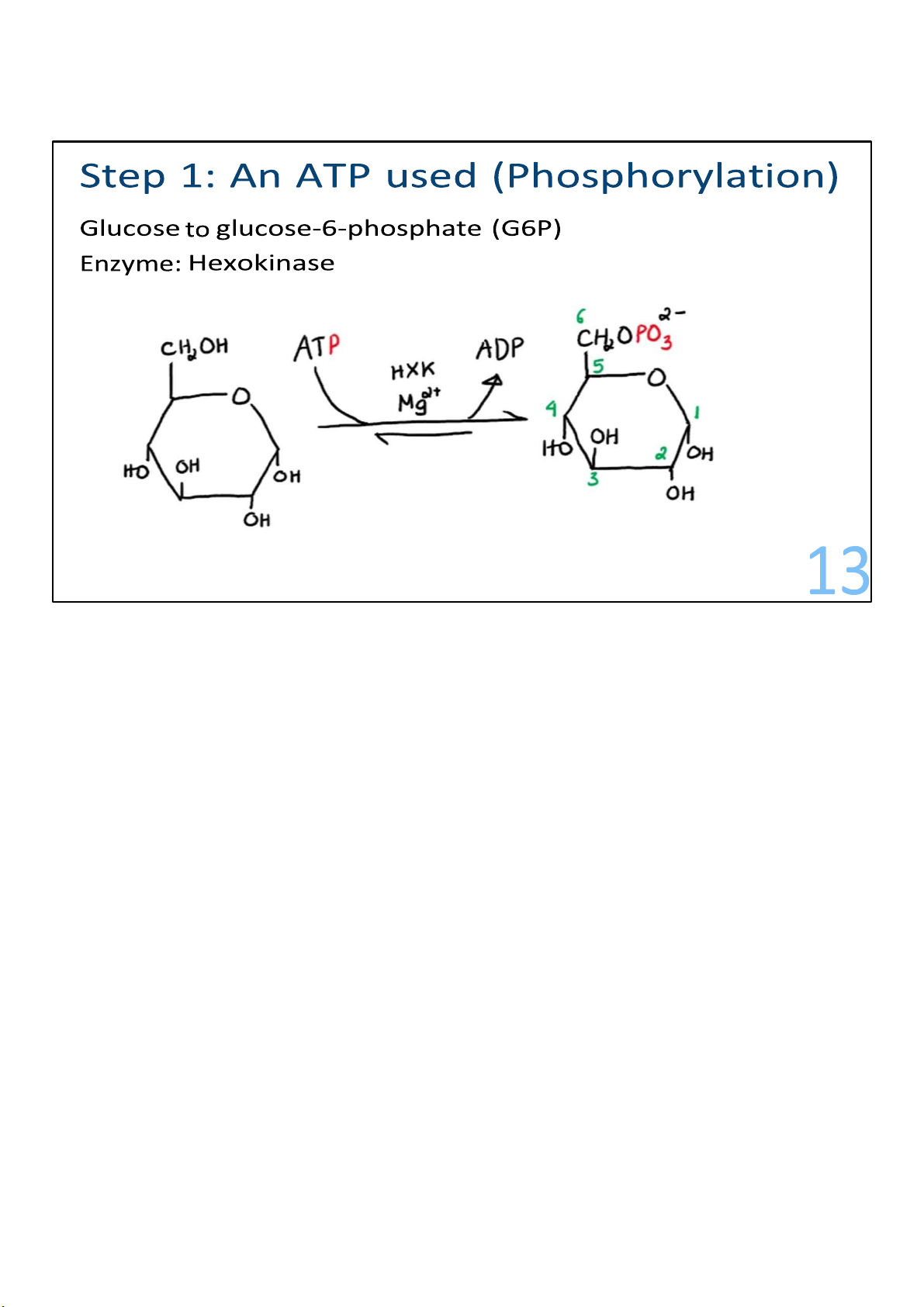
In the first step, a phosphate from ATP is transferred onto
carbon 6 of glucose to create a molecule known as glucose
6phosphate (G6P). The number 6 means the carbon where the
phosphate group interacts with. (Draw) I can call it
Phosphorylation, or in a more simple term, where an ATP is
used. Note that some of the titles I’ve got for the steps are not
by any mean official terminology or anything. I just put them
there in hope that they can make you understand what’s happening. 12 lOMoARcPSD|364 906 32
This requires ATP instead of producing it. To make ATP, we have
to use some ATP. Enzymes that put phosphates onto molecules 13 lOMoARcPSD|364 906 32
are kinases, and this one is called hexokinase. Adding a
phosphate onto glucose makes it negatively charged and
therefore makes it not able to easily exit cells on its own. 13 lOMoARcPSD|364 906 32 14
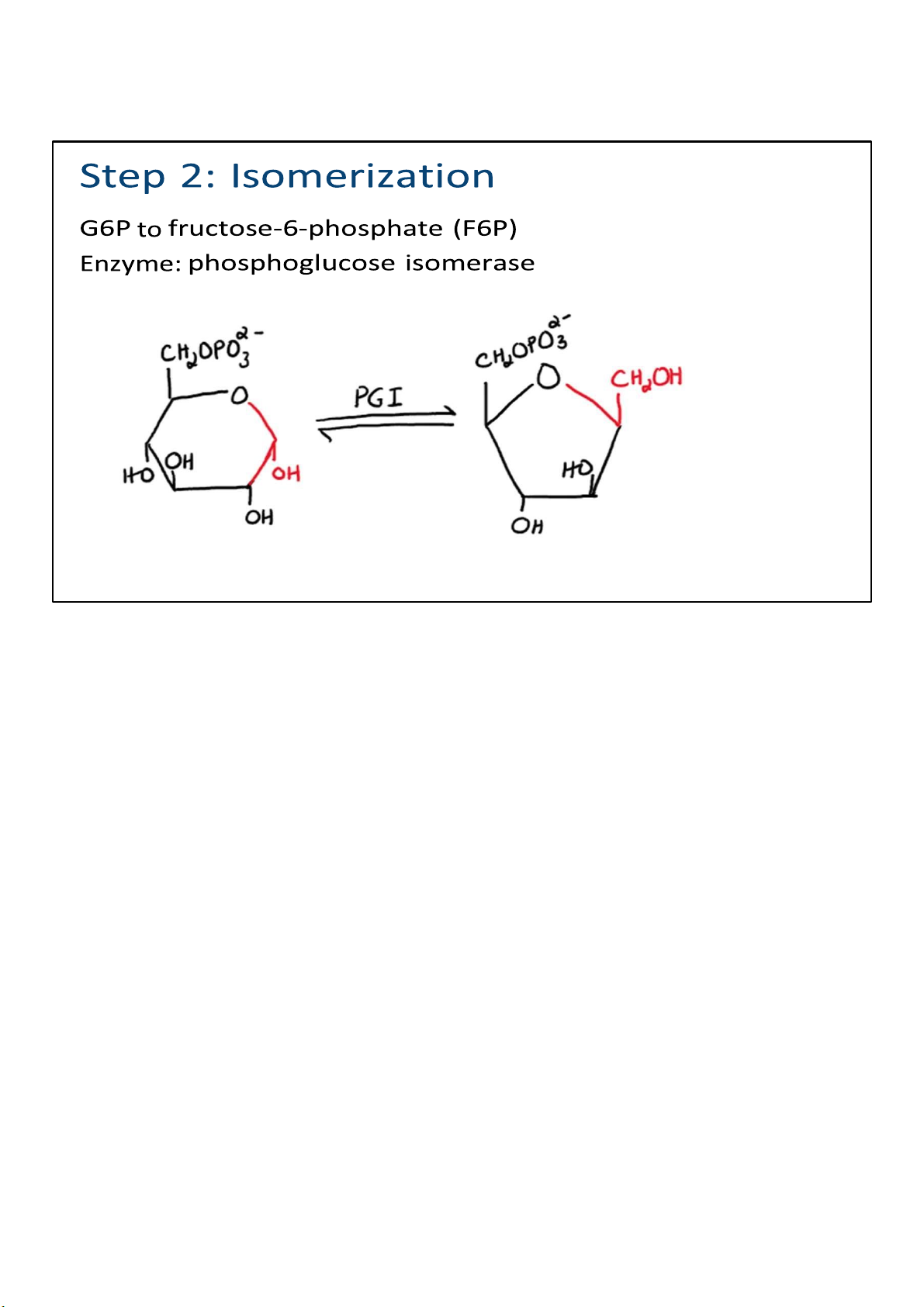
In step 2, there is a simple rearrangement of G6P. In other
words, nothing is added or taken away; the atoms in the G6P
are just rearranged to make fructose 6-phosphate (F6P). The
proper term would be isomerization, and of course it is catalyzed by an isomerase. 14 lOMoARcPSD|364 906 32
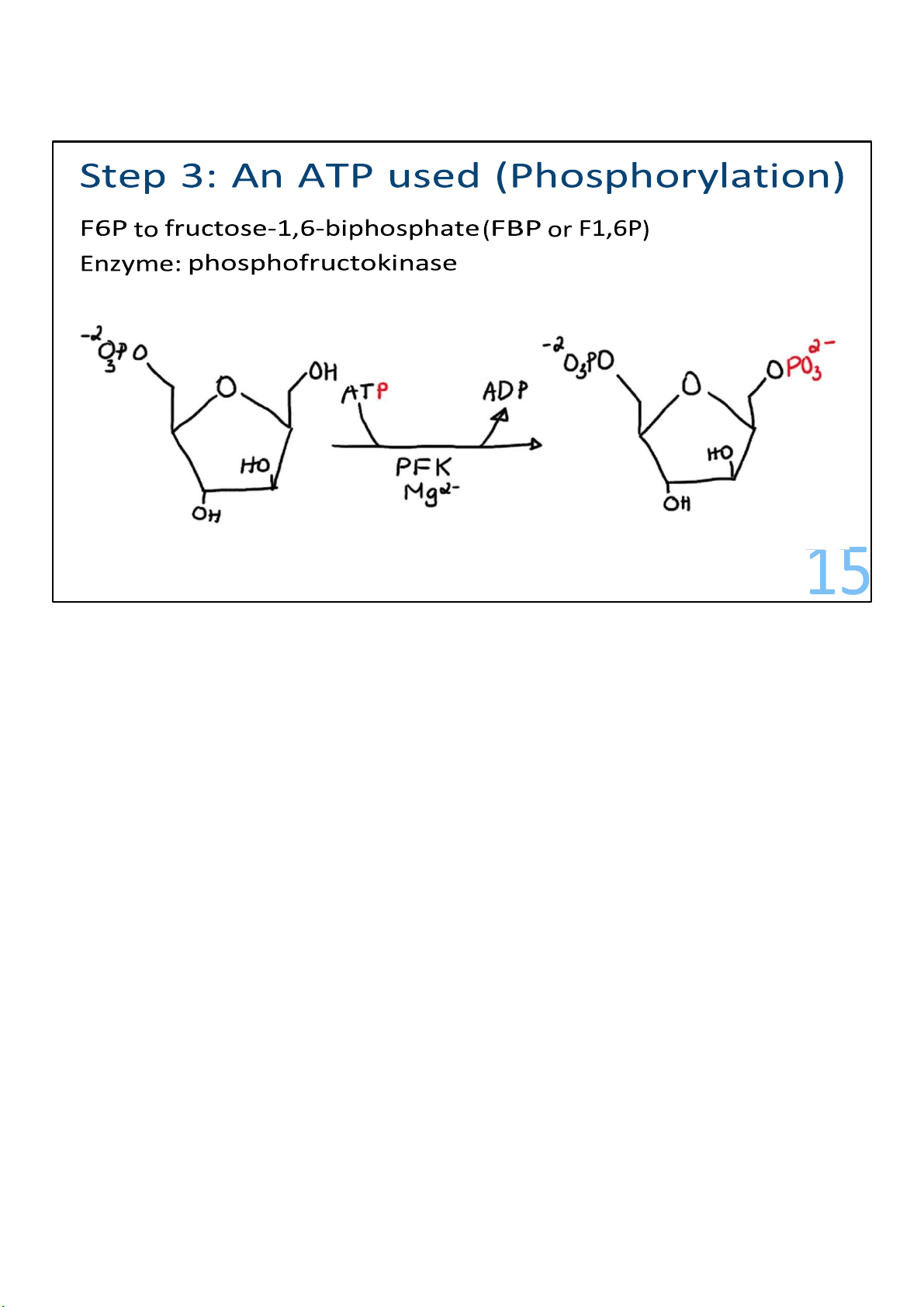
F6P is the substrate for reaction 3, a crucial control point for the pathway.
A second ATP is invested to add a second phosphate onto F6P,
this time at carbon 1, making fructose 1,6-bisphosphate
(F1,6BP). The enzyme catalyzing the reaction is called
phosphofructokinase (PFK). This enzyme is capable of
controlling the entire pathway. 15 lOMoARcPSD|364 906 32
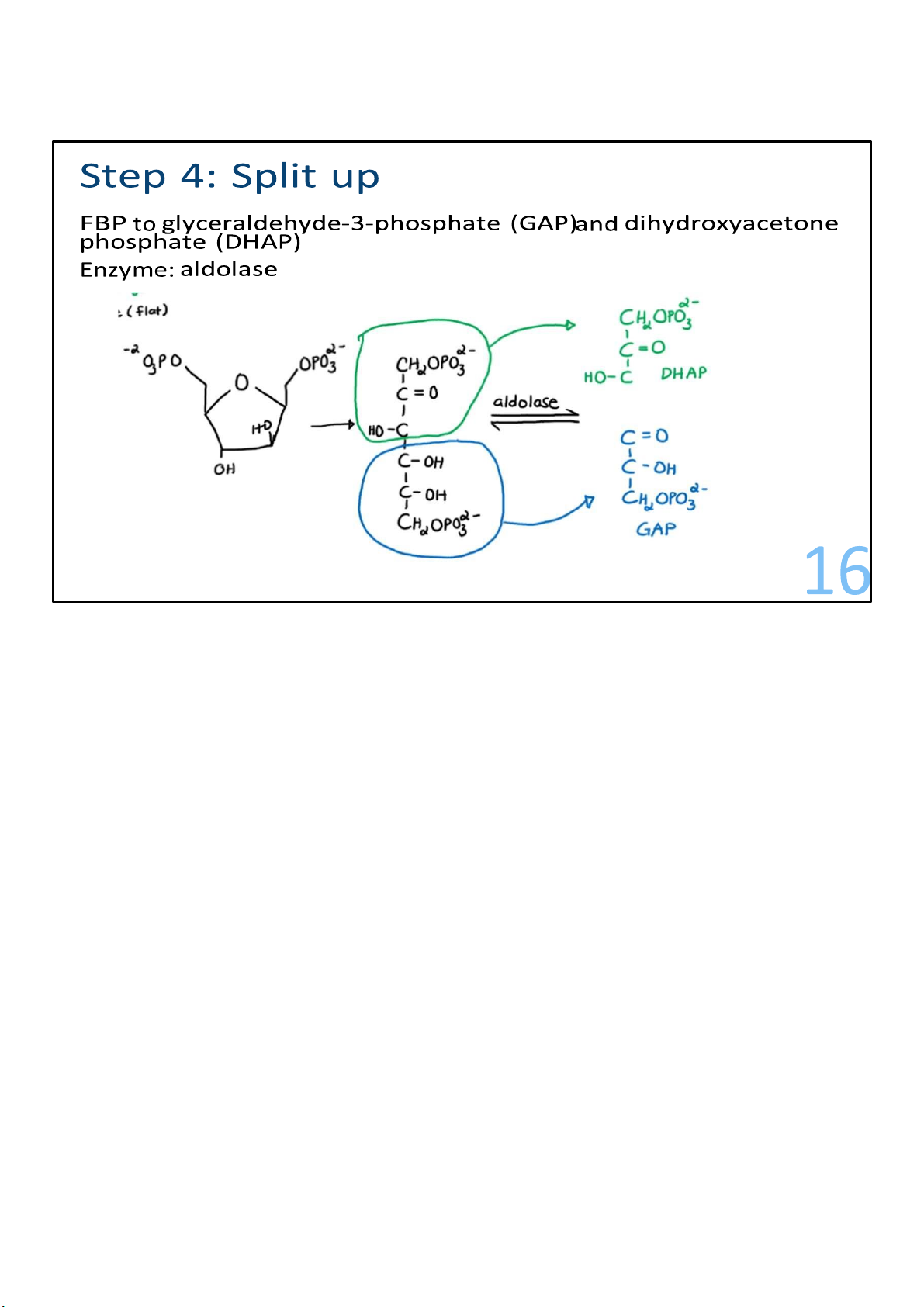
Catalyzed by the enzyme called aldolase, this step of glycolysis
splits F1,6-BP into 2 pieces of 3 carbons each. These 2 pieces
are not identical but instead are isomers, meaning that one of
them can be rearranged into the other. 16 lOMoARcPSD|364 906 32
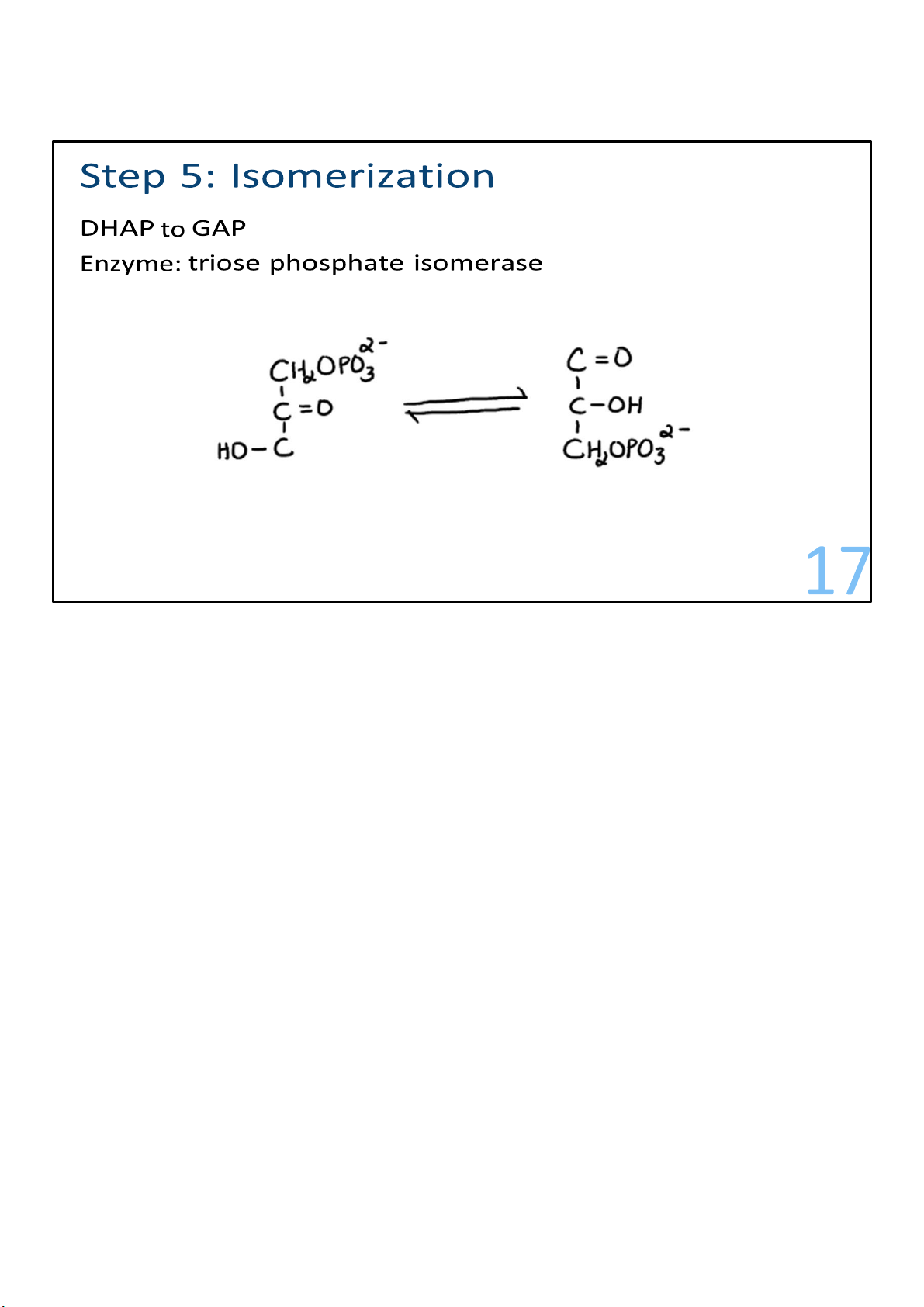
The piece called dihydroxyacetone phosphate (DHAP) gets
converted to its isomer, glyceraldehyde 3-phosphate (G3P). 17 lOMoARcPSD|364 906 32 18 lOMoARcPSD|364 906 32 Stage 2: Energy recovery Steps 6-10
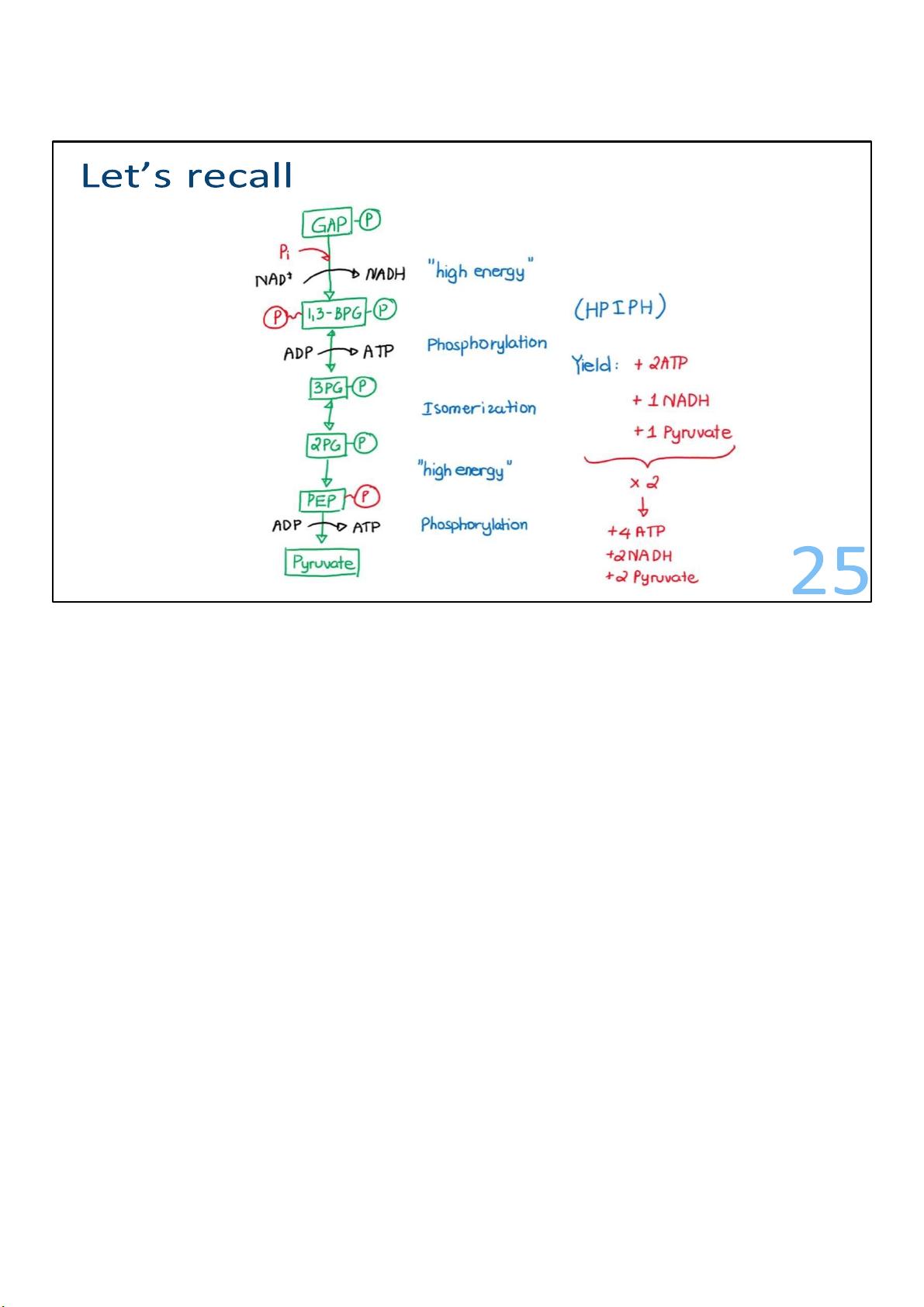
2 glyceraldehyde-3-phosphate to 2 pyruvate 19 19 lOMoARcPSD|364 906 32
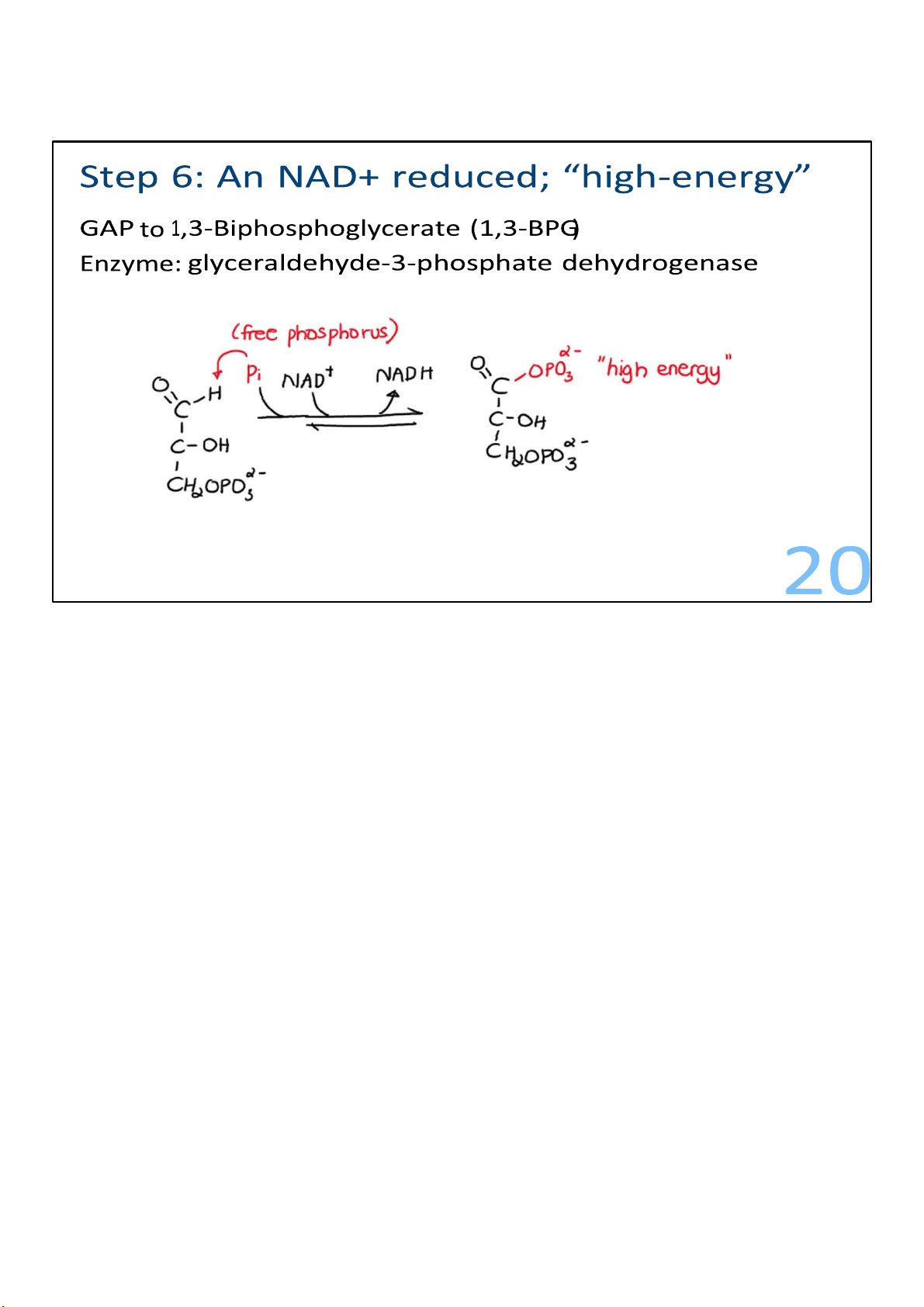
Glyceraldehyde 3-phosphates are converted to
1,3bisphosphoglyerates. This involves the addition of a
phosphate. For this to happen, the G3P aldehyde is converted
to an acid, which is an oxidation. And in that process, the
molecule being oxidized loses 2 electrons. It gives those
electrons to NAD+, which accepts them plus a proton and makes NADH. In the same 20 lOMoARcPSD|364 906 32
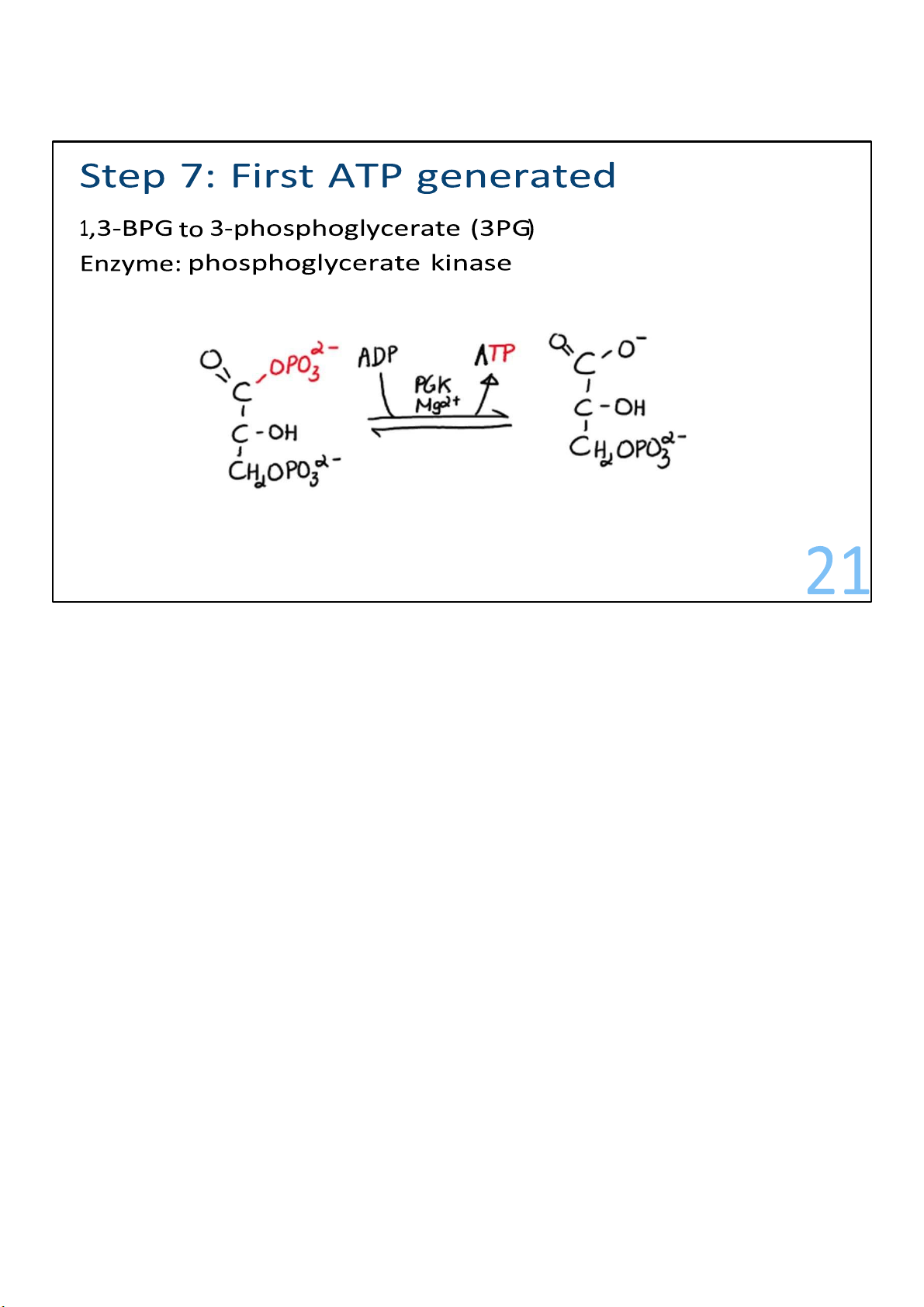
The formation of 1,3-bisphosphoglycerate kicks off the last
reactions of glycolysis, where ATP begins to get made! First, a
phosphate gets transferred away from each of the
1,3bisphosphoglycerates to ADPs, resulting in 2 ATPs! And 2
molecules called 3-phosphoglycerate are left behind. The
energy invested in steps 1 and 3 has been recovered. 21 lOMoARcPSD|364 906 32
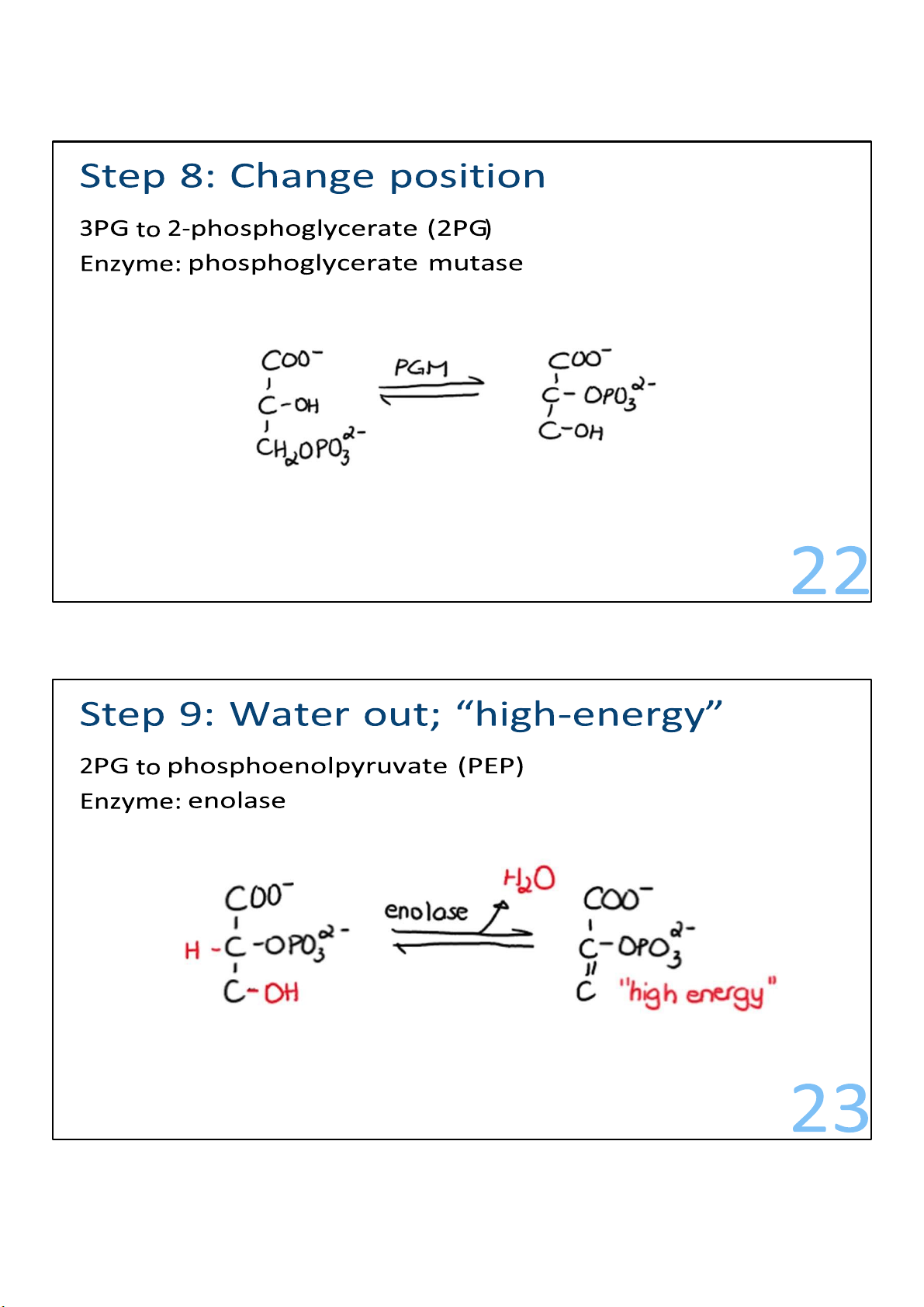
The molecules of 3-phosphoglycerate undergo rearrangement. 22 lOMoARcPSD|364 906 32
The molecules of 3-phosphoglycerate form a molecule named
phosphoenolpyruvate (PEP), which is the substrate for the
final step in glycolysis: the big bang.
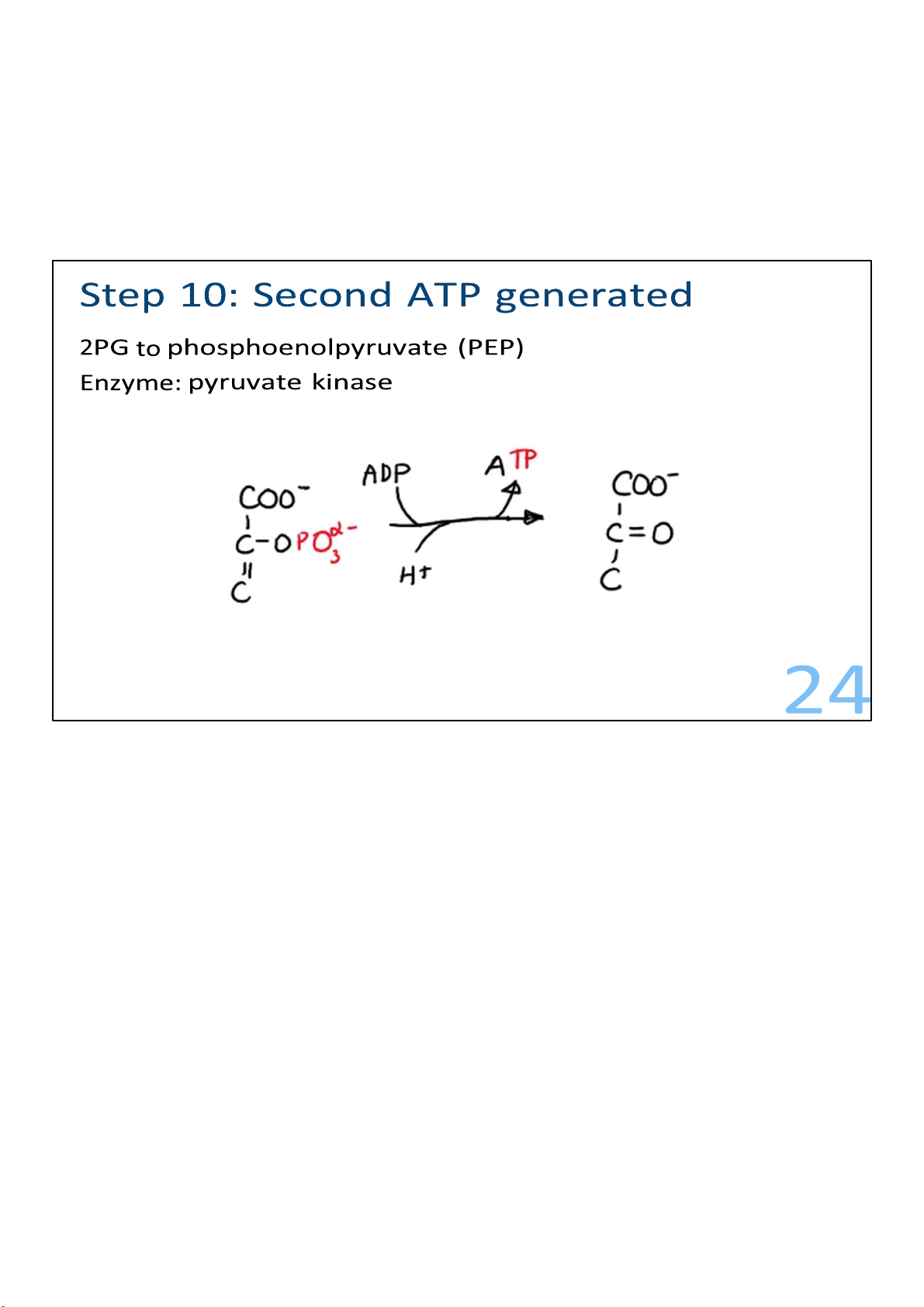
The big bang reaction, which is catalyzed by pyruvate kinase,
gets its nickname from a very large energy release that results.
In the reaction, 2 PEPs are converted to 2 pyruvates, and 2
ATPs are produced. There is enough energy released in the
reaction to almost produce 2 more ATPs, but not quite. Energy
that is released during reactions that is not captured or used is lost as heat. 23 lOMoARcPSD|364 906 32 24 lOMoARcPSD|364 906 32
The payoff, in terms of ATP, is double the investment. Regulating glycolysis
• Multiple steps / enzymes checkpoints to regulate
• Step 1: Feedback inhibition
High levels of product (G6P) inhibit enzyme HXK activity
• Step 3: Allosteric regulation ATP slows PFK down AMP speeds PFK up
F2,6-BP speeds PFK up (in response to insulin) 27 25 lOMoARcPSD|364 906 32
Having multiple steps also provides several points at which the
flow of intermediates through glycolysis can be adjusted.
Metabolic pathways must be regulated.
Step 1 of the pathway, catalyzed by the enzyme hexokinase,
makes G6P from glucose. However, the enzyme is inhibited by
high levels of the product, G6P. This feedback keeps cells from
piling up large amounts of phosphorylated glucose.
Step 3 of glycolysis—in which the enzyme phosphofructokinase
(PFK) adds a phosphate onto F6P, making F1,6-BP—is
allosterically regulated by 3 different molecules: ATP, a
lowenergy cousin called adenosine monophosphate (AMP),
and a double-phosphate molecule known as fructose
2,6bisphosphate (F2,6-BP). Allosteric regulating molecules bind to 26 lOMoARcPSD|364 906 32
an enzyme and alter its activity. When any of these
molecules binds to PFK, the resulting change in the
enzyme’s conformation dials the activity of the entire pathway up or down.
An abundance of ATP indicates that the cell has plenty of
energy, so when it binds to PFK, the enzyme’s activity
slows down. Large amounts of AMP, on the other hand,
signal low energy and a need for glycolysis to make ATP.
The binding of AMP to PFK increases PFK activity.
F2,6-BP is made in response to insulin, which stimulates
cells to take up glucose following a meal. When this
happens, cells must run glycolysis to break it down. So,
F2,6-BP increases PFK enzyme activity and speeds up glycolysis. 27 lOMoARcPSD|364 906 32 28 lOMoARcPSD|364 906 32 Glycolysis and other pathways
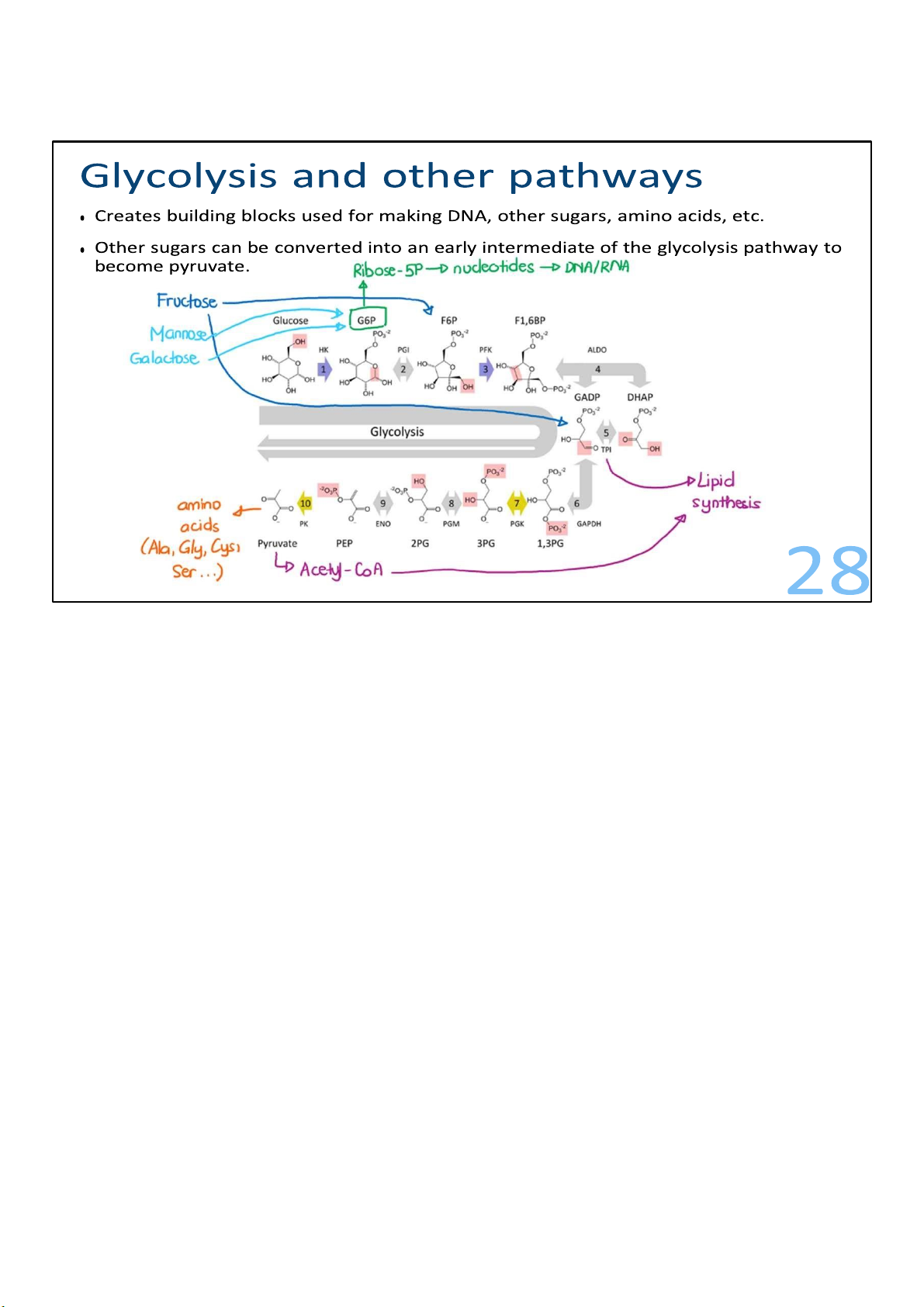
• Other sugars can be converted into an early intermediate of the glycolysis pathway to become pyruvate. 29
In addition to glucose, glycolysis is also important for the
metabolism of other sugars, such as fructose, mannose, and
galactose. Each of these sugars is readily converted into an
early intermediate of the glycolysis pathway and can get made into pyruvate. 29 lOMoARcPSD|364 906 32 What happens after?
• With oxygen: Citric acid cycle • No oxygen:
• Lactic fermentation (yogurt, sauerkraut, in our muscle in vigorous exercise)
• Alcohol fermentation (wine, beer) 30
Just as there is more than one way to enter the glycolysis
pathway, there are also alternatives about what happens at the
end. In particular, when there is no oxygen, there are
alternative exits from glycolysis.
One such exit is lactic fermentation. It produces lactic acid,
which is used to make foods like yogurt or sauerkraut. It is also
the path used by our muscle cells during prolonged, vigorous
exercise, when the oxygen supply to muscles is unable to keep up with the demand.
A second fermentation pathway that other yeasts and bacteria
run in the absence of oxygen produces alcohol from pyruvate.
This is the exit from glycolysis that wine and beer makers
depend on. In this 2-step process, pyruvate is first converted to 30 lOMoARcPSD|364 906 32
acetaldehyde, and carbon dioxide is released in the
process. The acetaldehyde is then turned into alcohol
using electrons from NADH, thus regenerating NAD+. 30 lOMoARcPSD|364 906 32 Citric acid cycle 31 Where metabolism meets
Glycolysis generates some ATP, but its main role is to deliver molecules that can be further
oxidized to extract more energy. This is where the citric acid cycle comes in, providing a second
stage for the efficient production of energy from food.
Sugars, fats, and amino acids all converge on a single
oxygendependent metabolic pathway known as the citric acid
cycle, or the Krebs cycle. Even more than glycolysis, the citric
acid cycle is a central metabolic pathway where everything
comes together. And this one pathway is used by every cell,
from the cells in bacteria to amoebas to you. It’s a cycle
• Also called Krebs cycle or tricarboxylic acid (TCA) cycle
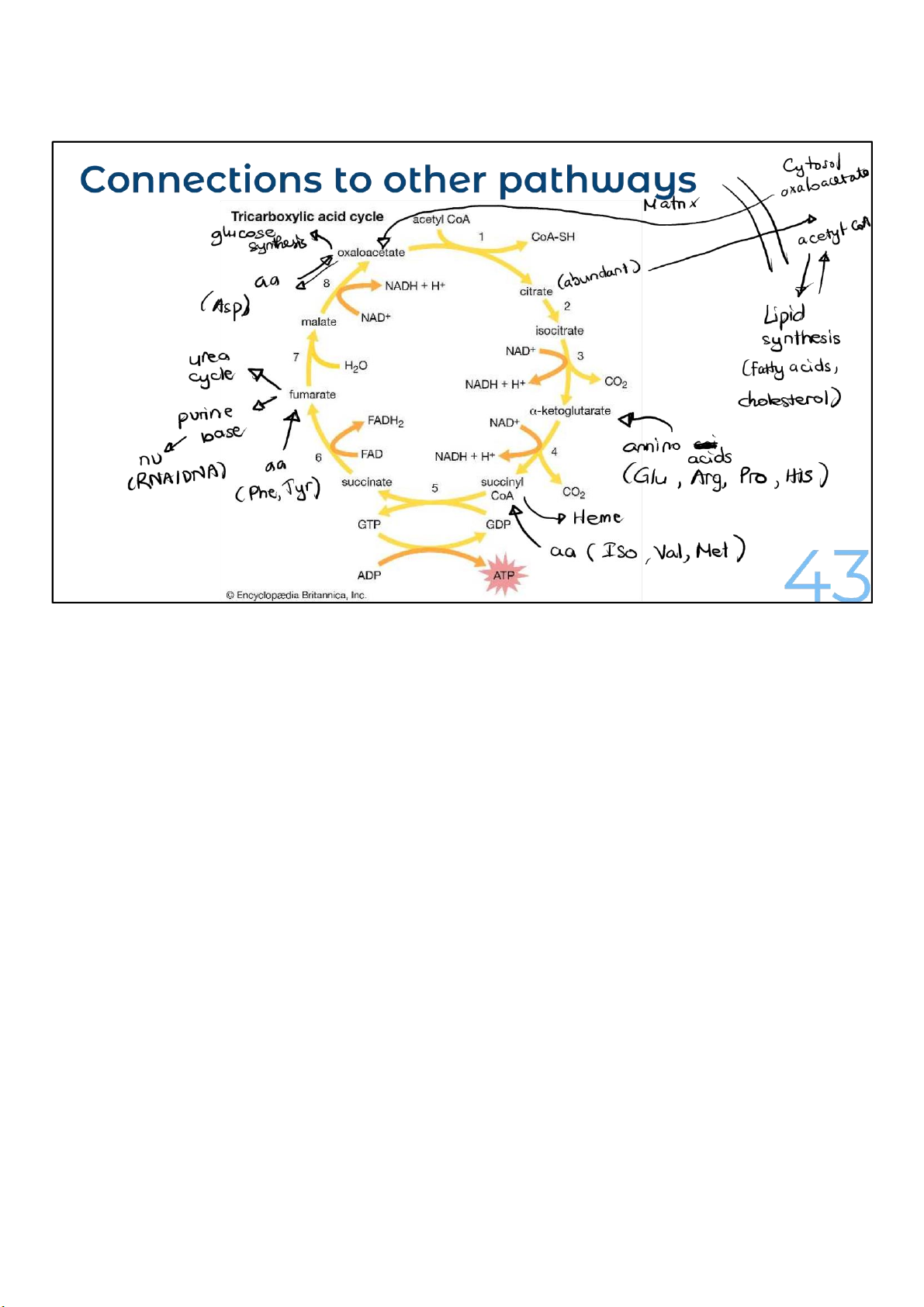
• Extract energy & provides intermediates for other pathway (like glycolysis)
• Return to its starting point (oxaloacetate)
• 1 + 8 steps, including 8 in the cycle 31 lOMoARcPSD|364 906 32 32
Like glycolysis, the citric acid cycle extracts energy and provides
intermediates for other pathways. As a circle, the citric acid
cycle returns to its starting point. And molecules from other
pathways can enter it at multiple points and leave as needed.
Just as in glycolysis, the oxidation of the intermediates in the
citric acid cycle is coupled with energy capture, both in
phosphate bonds and in molecules of the activated carriers, NADH and FADH2.
Acetyl-CoAs can enter the citric acid cycle directly. But
pyruvate made in glycolysis needs an extra step to get
converted to acetyl-CoA, and then it, too, can enter. To start 32 lOMoARcPSD|364 906 32
the process of entering the pathway, pyruvate needs to move
from the cytosol (the cytoplasm), where glycolysis produced it,
to an innermost section of mitochondria called the (mitochondrial) matrix.
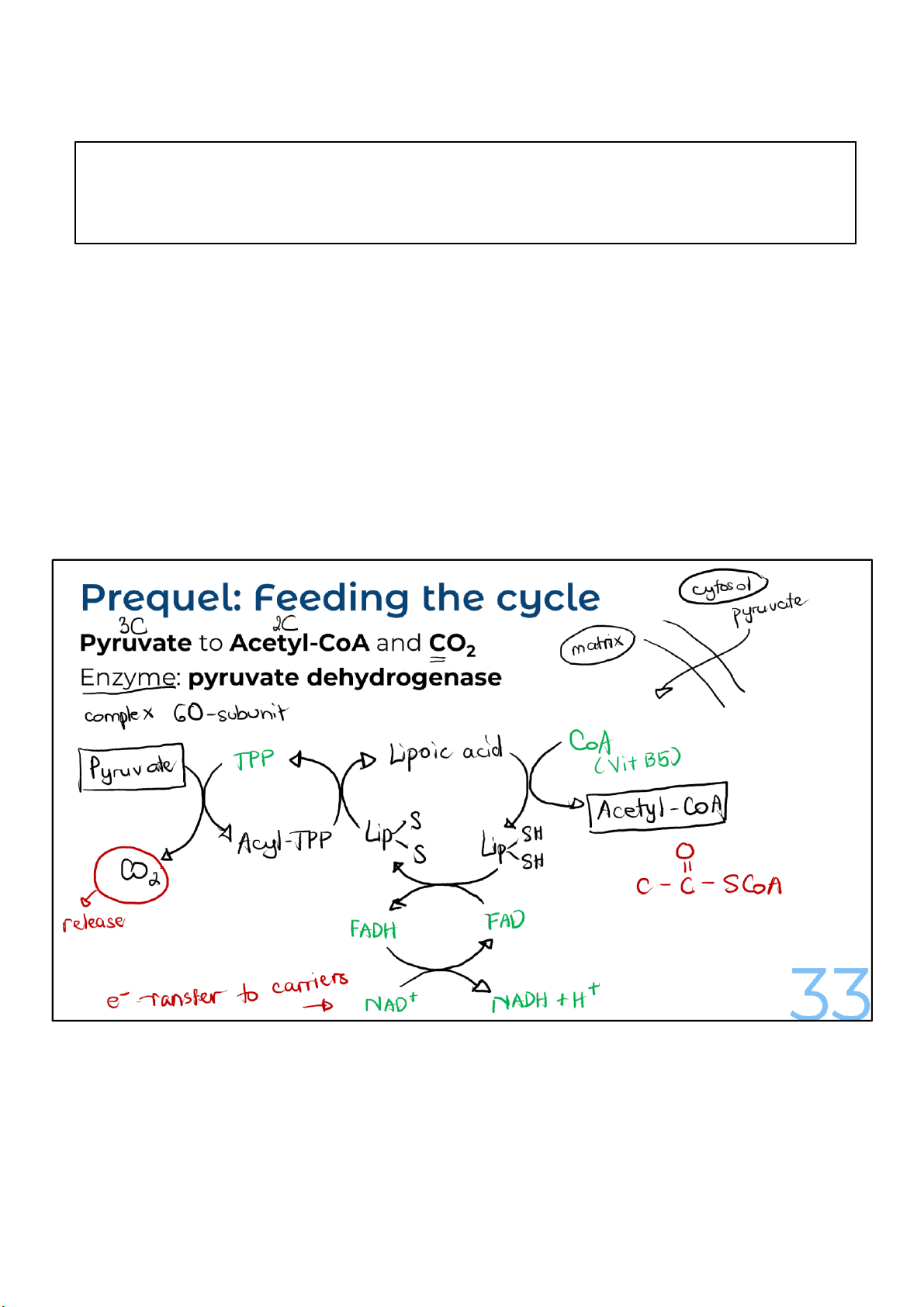
To get converted to acetyl-CoA, pyruvate is first transported
into mitochondria, where an enzyme complex called pyruvate
dehydrogenase acts on it. Remember, the conversion of
pyruvate to acetyl-CoA is necessary for the entry of pyruvate into the cycle.
The reason pyruvate dehydrogenase is called a complex is
because it is not just one enzyme, but several enzymes that 33 lOMoARcPSD|364 906 32
catalyze the multiple reactions necessary for converting
pyruvate to acetyl-CoA. The enzyme complex uses 5
different cofactor molecules that act as helpers for the
reactions. These cofactors include lipoic acid, coenzyme
A, thiamine pyrophosphate, FAD, and NAD+. The last 4
of these molecules are either vitamins or are derived from them. 33 lOMoARcPSD|364 906 32
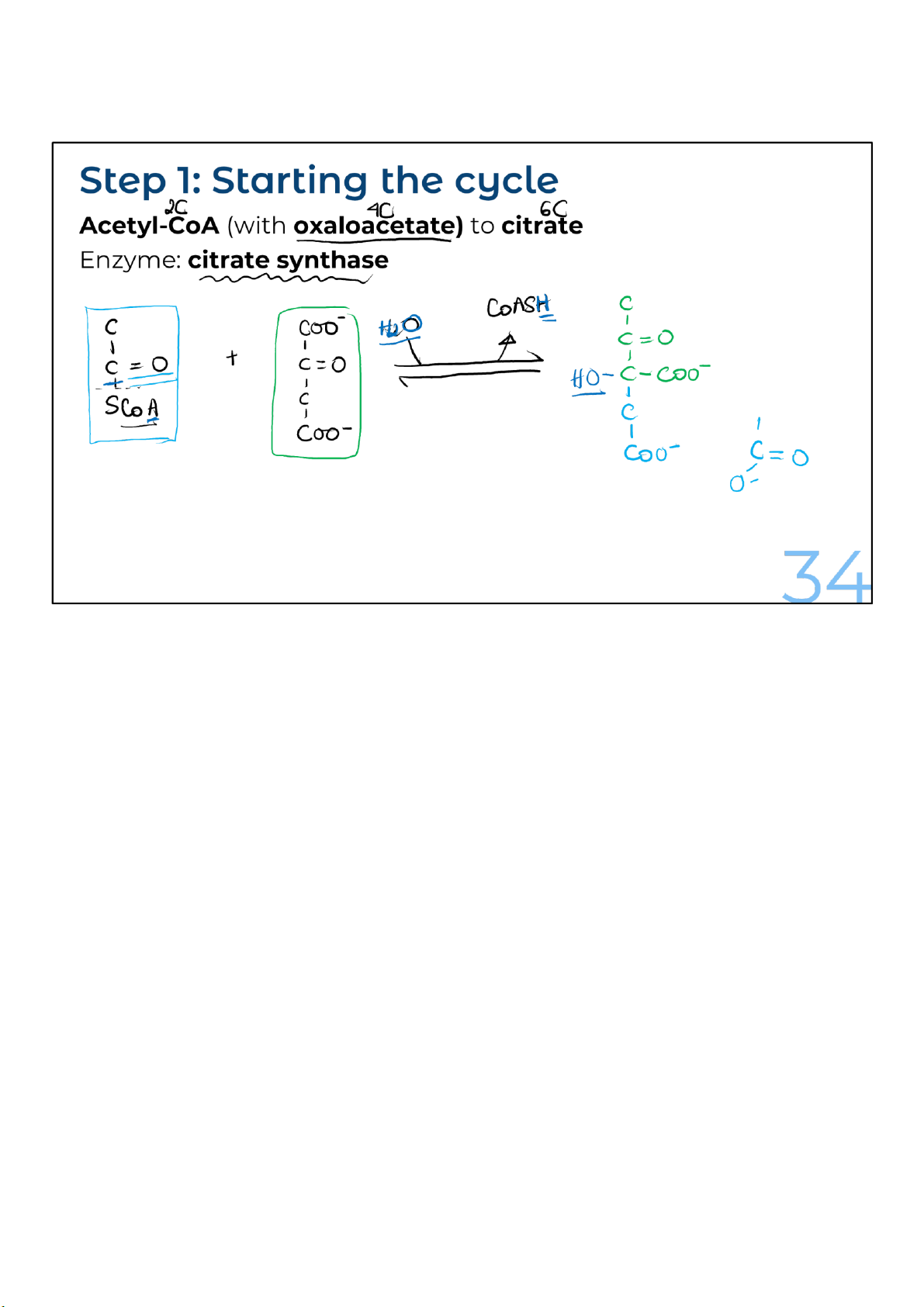
Acetyl-CoA transfers its 2-carbon acetyl group to a 4-carbon
molecule of oxaloacetate, which comes from the “end” of the
cycle. The 4-carbon oxaloacetate and the 2-carbon acetyl
group combine to produce a 6-carbon molecule called citrate,
which is the name for citric acid when it’s ionized.
This reaction to create citrate is very favorable because
breaking the bond between the acetyl group and coenzyme A
releases a lot of energy. This turns out to be important when
the circle is completed to make oxaloacetate. 34 lOMoARcPSD|364 906 32
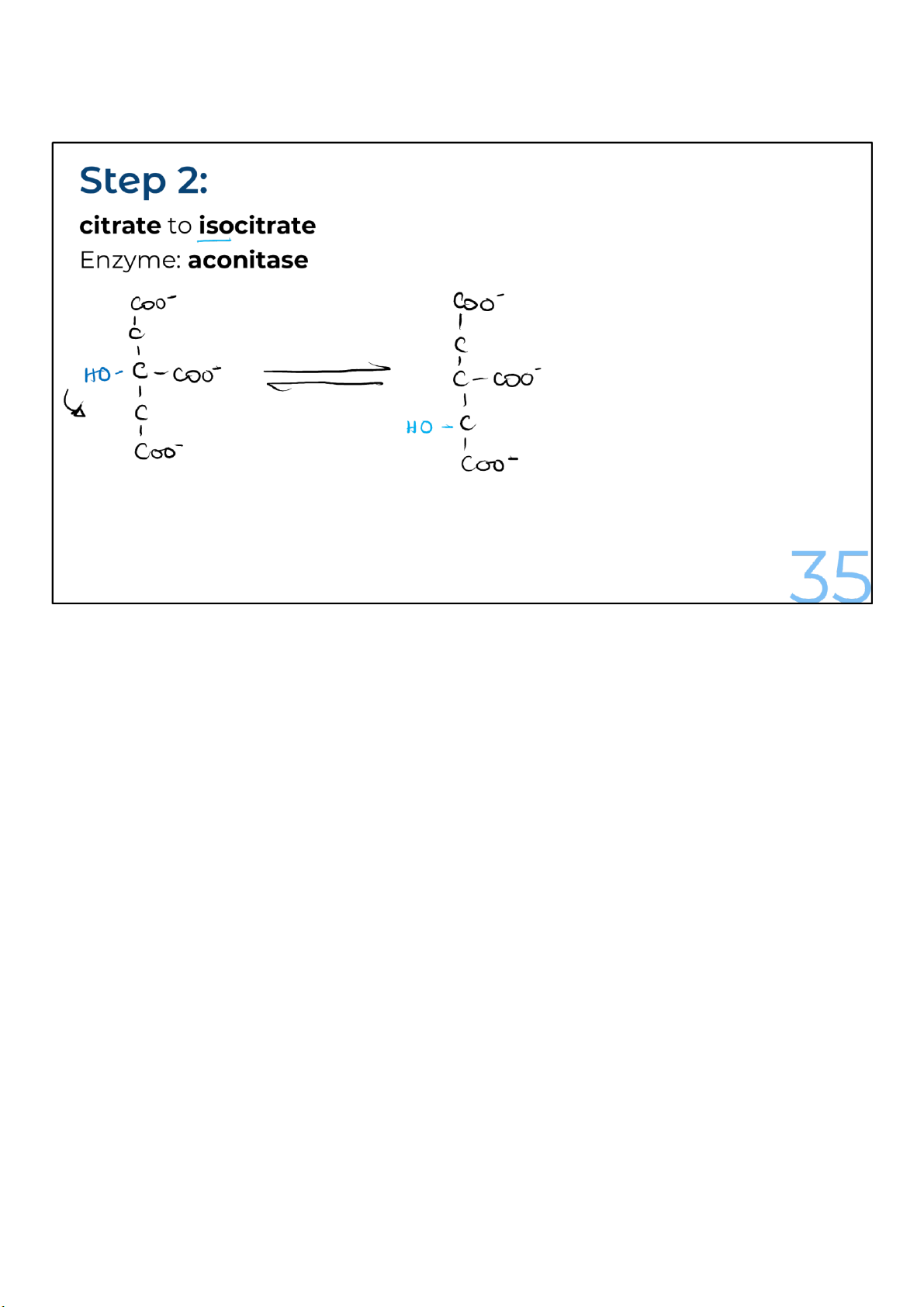
Citrate is rearranged to form its isomer, isocitrate. The enzyme
catalyzing the reaction, aconitase, is the site of action of a
poisonous compound known as fluoroacetate, which cells
readily convert into fluorocitrate. That’s a big problem
because fluorocitrate is a potent inhibitor of the aconitase enzyme.
Inhibiting this enzyme blocks the entire citric acid cycle. 35 lOMoARcPSD|364 906 32
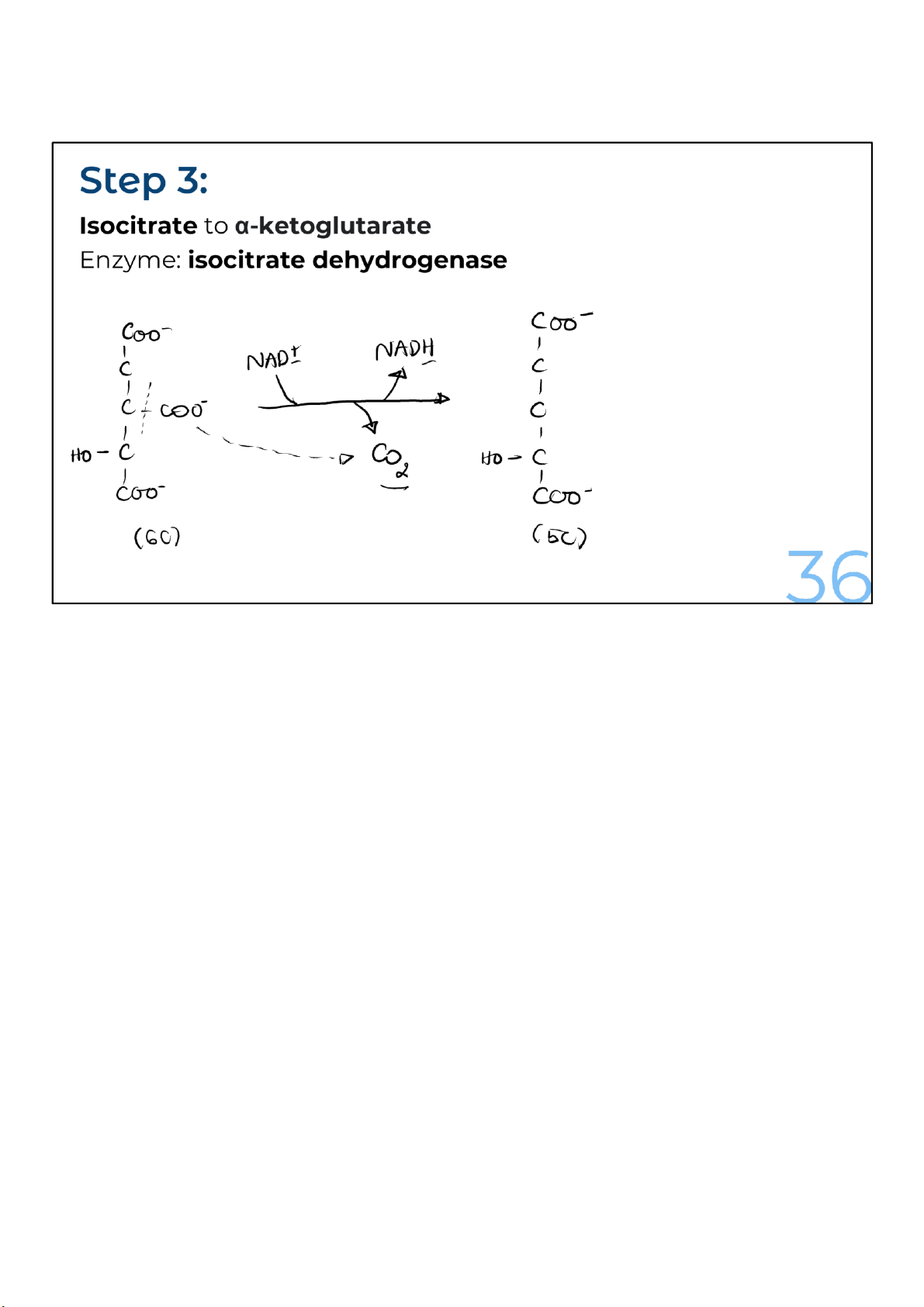
The isocitrate loses a carbon dioxide and a pair of electrons in
a process called oxidative decarboxylation. The loss of a
carbon as carbon dioxide converts the 6-carbon isocitrate into
a 5carbon molecule, alpha-ketoglutarate. Meanwhile, the
electrons from isocitrate get transferred to NAD+, making
NADH. Remember, the oxidation of isocitrate (the loss of
electrons) causes NAD+ to be reduced to NADH. 36 lOMoARcPSD|364 906 32
The same thing happens to alpha-ketoglutarate as another
carbon is lost as carbon dioxide. Again, electrons are given
away, and they combine with NAD+ to form an NADH. The
product of the reaction, a 4-carbon compound, links to a CoA,
forming succinyl-CoA. The enzyme here, alpha-ketoglutarate
dehydrogenase, is closely related to pyruvate dehydrogenase;
it uses the same 5 coenzymes and has a similar reaction mechanism. 37 lOMoARcPSD|364 906 32
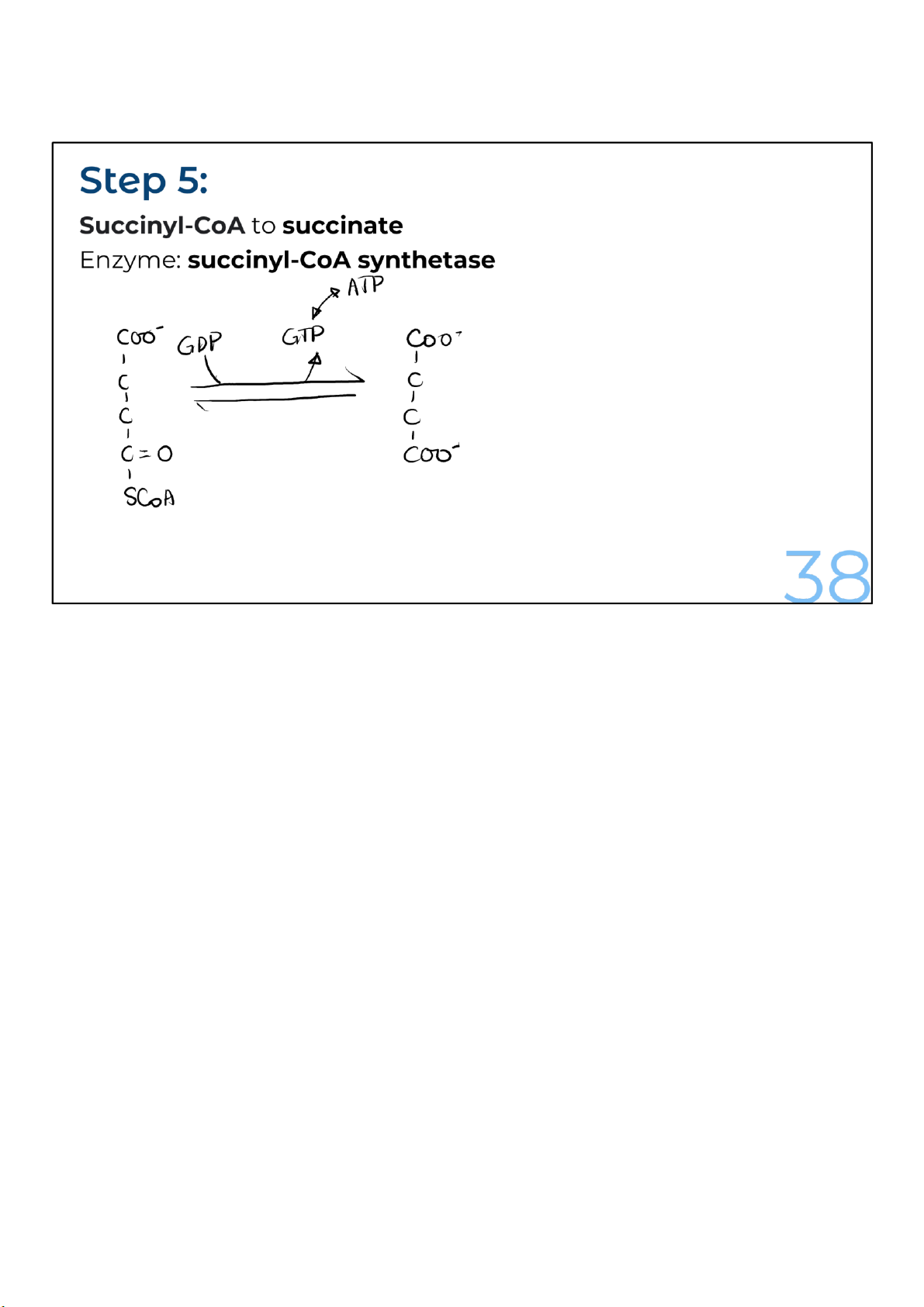
The coenzyme A is removed from succinyl-CoA to produce succinate.
The removal of the CoA from succinyl-CoA is also a very
energetically favorable reaction, just like when the CoA was
released from acetyl-CoA in the first reaction. That’s because
the bond between succinate and coenzyme A is a high-energy
one. The energy released is used to convert guanosine
diphosphate (GDP) to guanosine triphosphate (GTP). The
energy in GTP is equivalent to that in ATP. 38 lOMoARcPSD|364 906 32
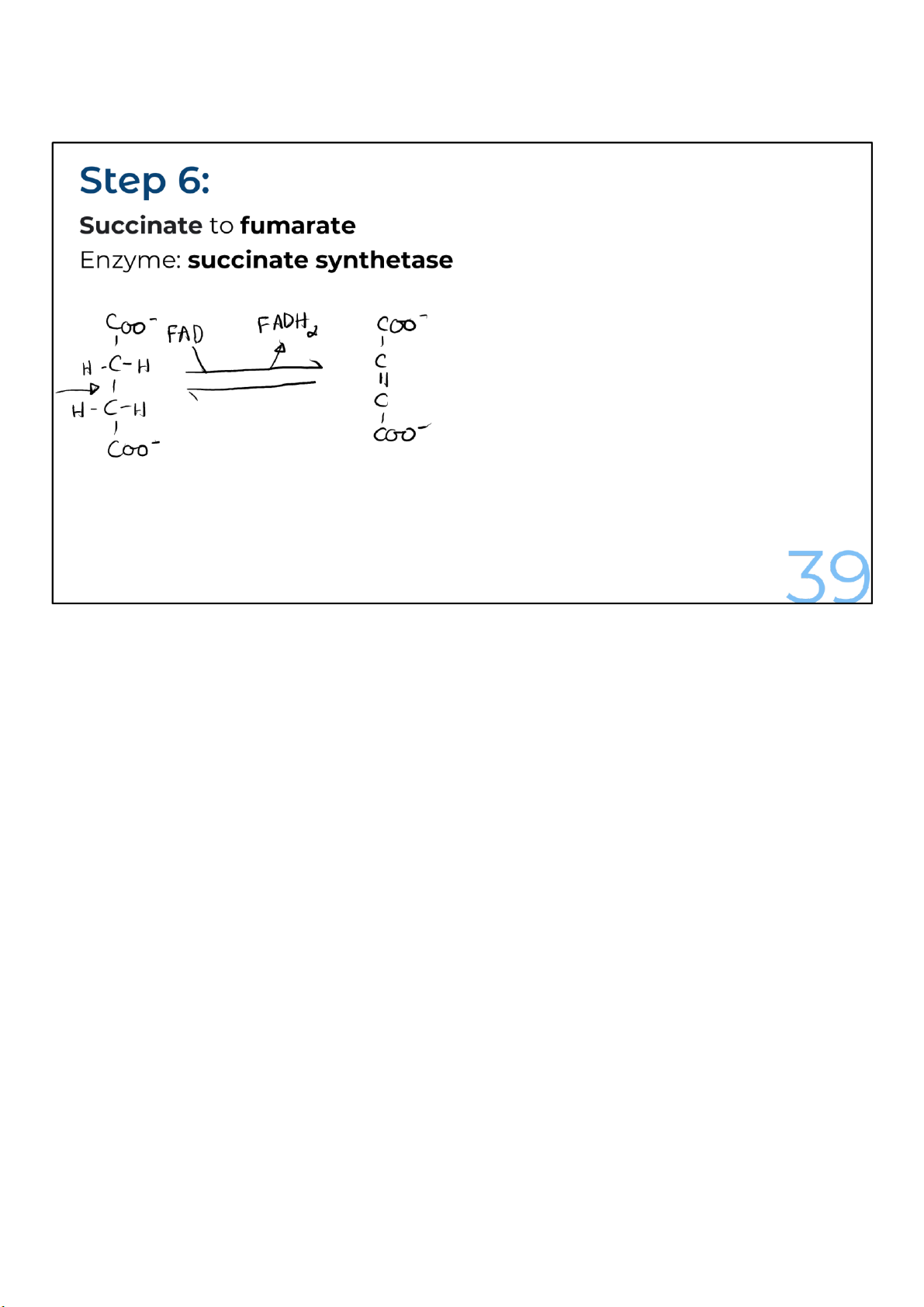
The remaining reactions in the cycle are focused on recreating
the oxaloacetate to complete the circle. In the first of these
reactions, succinate is oxidized to form fumarate by the
enzyme succinate dehydrogenase, which gives up electrons
and protons, which are accepted by FAD to make FADH2. 39 lOMoARcPSD|364 906 32
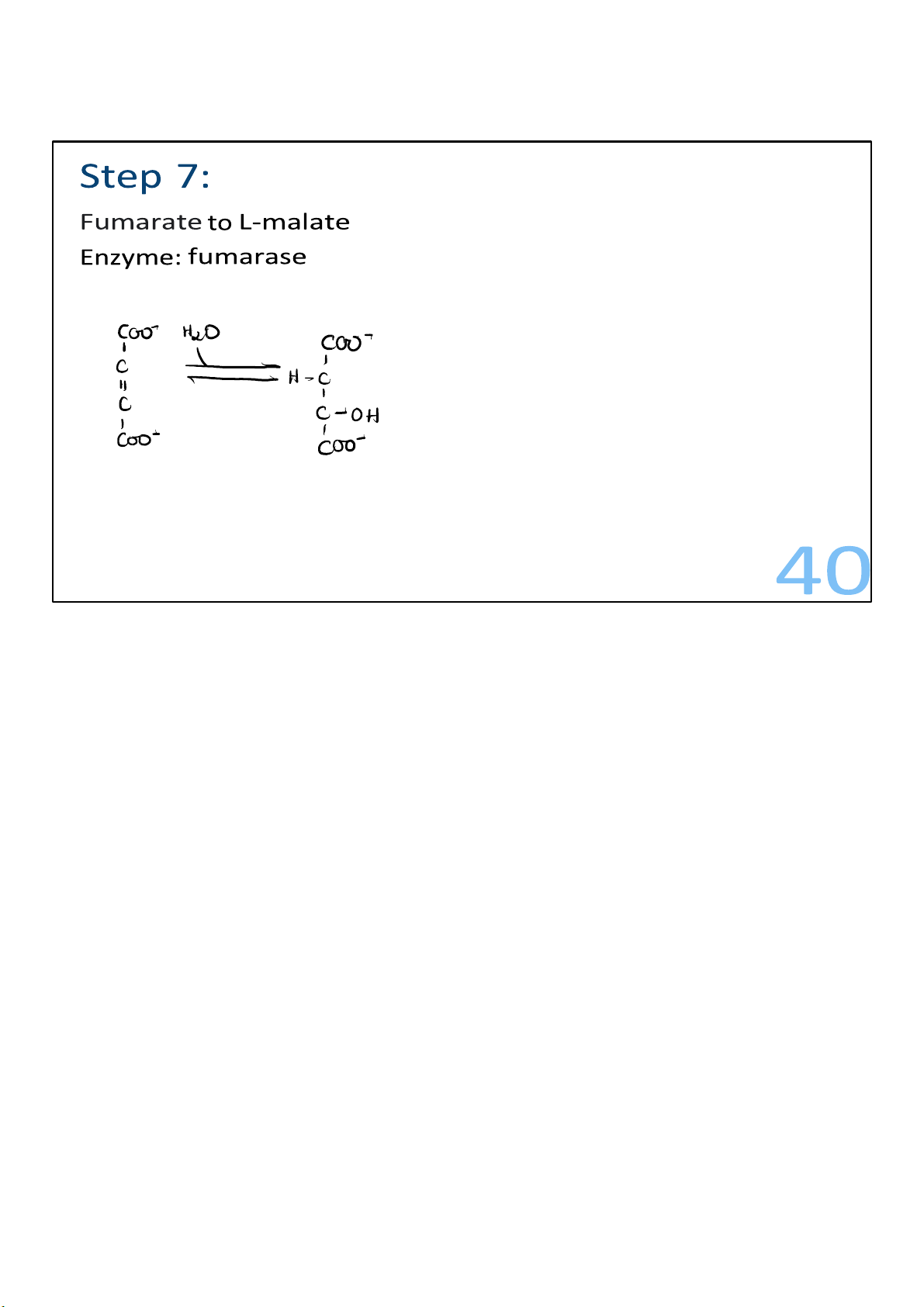
Water is added across the double bond of fumarate to form the molecule malate. 40 lOMoARcPSD|364 906 32
Malate is oxidized to oxaloacetate and, just as in many
oxidation reactions, NADH is produced. The reaction,
catalyzed by malate dehydrogenase, is notable for going
backward under standard conditions.
Fortunately, though, the cell is not under standard conditions.
The product gets removed, and the reaction gets pulled
forward by the citrate synthase reaction, which follows in the
next round of step 1. This reaction is energetically favored and
removes the product, oxaloacetate, as it reacts with acetyl-
CoA to form citrate. With oxaloacetate being quickly removed
to make citrate, the production of oxaloacetate can proceed. 41 lOMoARcPSD|364 906 32
With that, we’ve traveled around the cycle and returned to
where oxaloacetate reacts with acetyl-CoA to make citrate. 42 lOMoARcPSD|364 906 32
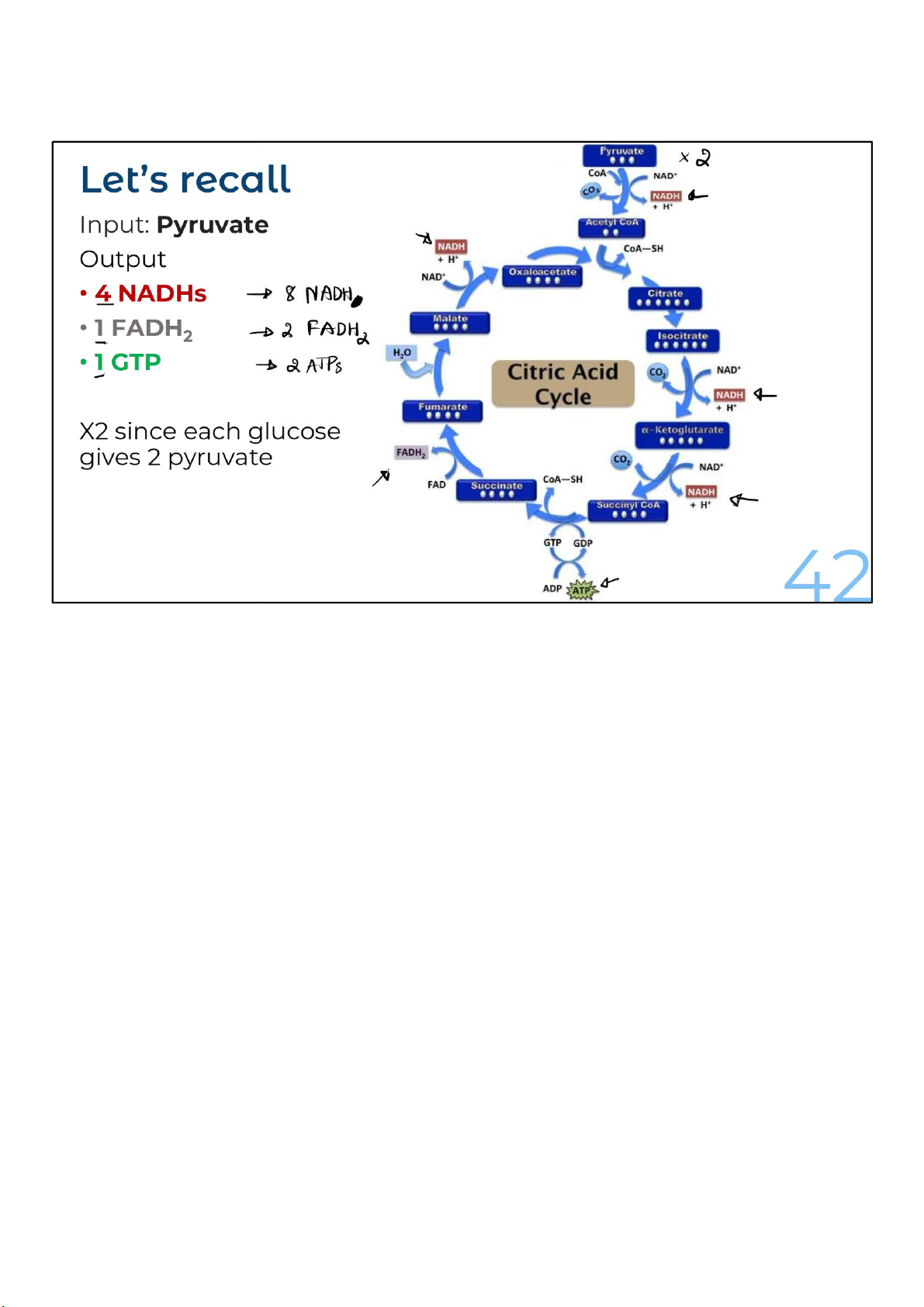
There are 2 turns of the cycle per glucose molecule
being oxidized. That’s because each glucose gives rise to
2 pyruvates, which in turn gives rise to 2 acetyl-CoAs. 41 lOMoARcPSD|364 906 32
The amount of energy that has been extracted through the
oxidative reactions of the citric acid cycle for the 2 acetyl-CoAs
is 6 NADHs, 2 FADH2s, and 2 molecules of GTP. And if pyruvate
was the source for the acetyl-CoAs, one NADH for each
pyruvate was received from glucose for a total of 8 NADHs, 2
FADH2s, and 2 molecules of GTP. 42 lOMoARcPSD|364 906 32 43 lOMoARcPSD|364 906 32 Electron Transport Chain
& Oxidative Phosphorylation
The final phase in energy harvesting 44
This is the final phase in energy harvesting, which comes after the citric acid cycle and fatty acid
oxidation (which you will learn in later lectures) and takes place in the innermost, liquid-filled region of
mitochondria called the matrix. This is why we need to build some foundation on the structure of
mitochondria, the power house of the cell. 44 lOMoARcPSD|364 906 32 The Structure of Mitochondria 45 45 lOMoARcPSD|364 906 32
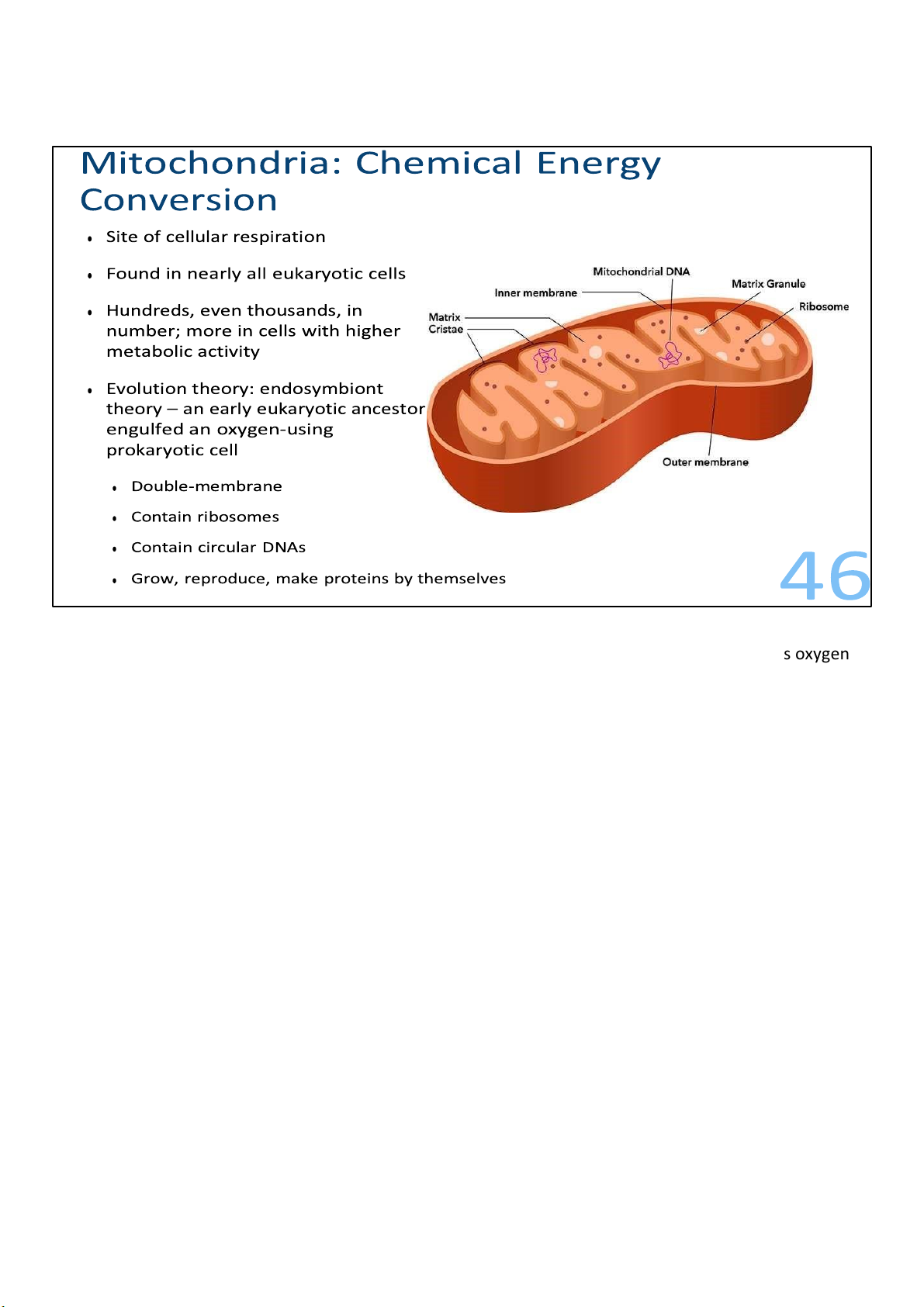
As mentioned, mitochondria are the sites of cellular respiration, the metabolic process that uses oxygen
to drive the generation of ATP, in contrast to fermentation, which does not involve oxygen.
They’re found in nearly all eukaryotic cells, including plants, animals, fungi, and most unicellular eukaryotes.
Although a number of cells have a single large mitochondrion, which is the singular form of
mitochondria, most of the time a cell has hundreds or even thousands of mitochondria; the number
correlates with the cell’s level of metabolic activity. That is, the more energy-consuming a cell is, the
more mitochondria it has. For example, cells that move or contract have proportionally more
mitochondria per volume than less active cells.
A pretty interesting theory about the origin of mitochondria, and also its counterpart chloroplasts, is the
endosymbiont theory. This theory states that an early ancestor of eukaryotic cells engulfed an
oxygenusing nonphotosynthetic prokaryotic cell. Eventually, they formed a relationship, or an
endosymbiont, meaning a cell living within another cell, a mutualism, meaning both cells benefit from
this arrangement. The host cell had a power house for much more efficient energy harvesting, and the
symbiont received protection from the hostile outside environment. Over the course of evolution, the
host cell and its endosymbiont merged into a single organism, a eukaryotic cell with a mitochondrion.
There are many evidence to prove this theory, and in fact, it’s a widely accepted theory. Firstly, it’s
consistent with many structural features of mitochondria, such as their double membrane while other
organelles are bounded by a single membrane. They also contain their own ribosomes, the assembly to 46 lOMoARcPSD|364 906 32
produce polypeptide chains, and circular DNA molecules – like bacterial chromosomes. And the DNAs
actually program the synthesis of some proteins they need for the respiration. They also grow and 47 lOMoARcPSD|364 906 32
reproduce on their own, and move around, changing their shapes, fusing, dividing within the cell. 46 lOMoARcPSD|364 906 32 – 10
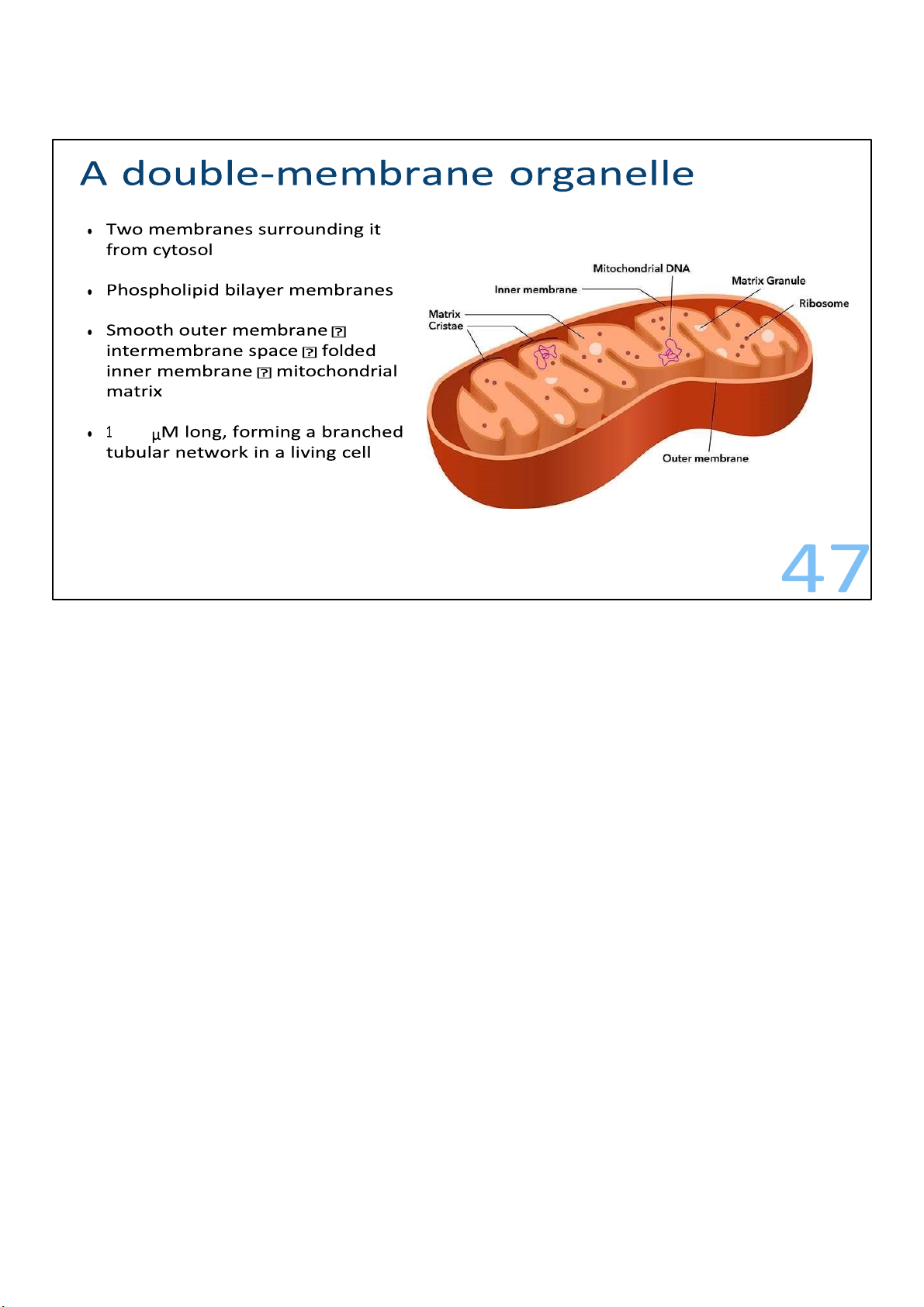
As I’ve said, mitochondria have two membranes surrounding them. Each of the membrane is a
phospholipid bilayer with some embedded proteins.
The outer membrane is smooth, but the inner is convoluted, with infoldings called cristae. The
membranes divide the mitochondria into 2 parts, the intermembrane space within the outer and inner,
and the compartment enclosed by the inner called the mitochondrial matrix, or matrix for short. With its
highly folded surfaces, the cristae give the inner membrane a larger surface area, thus enhancing the
productivity of cellular respiration.
All of these structure fits within a length of 1 to 10 micrometres. As the mitochondria move and fuse and
divide in the cell, they form a branched tubular network. 47 lOMoARcPSD|364 906 32
Electron Transport Chain (ETC)
Energy released helps pump out protons, creating a gradient 4 steps of moving electrons 48
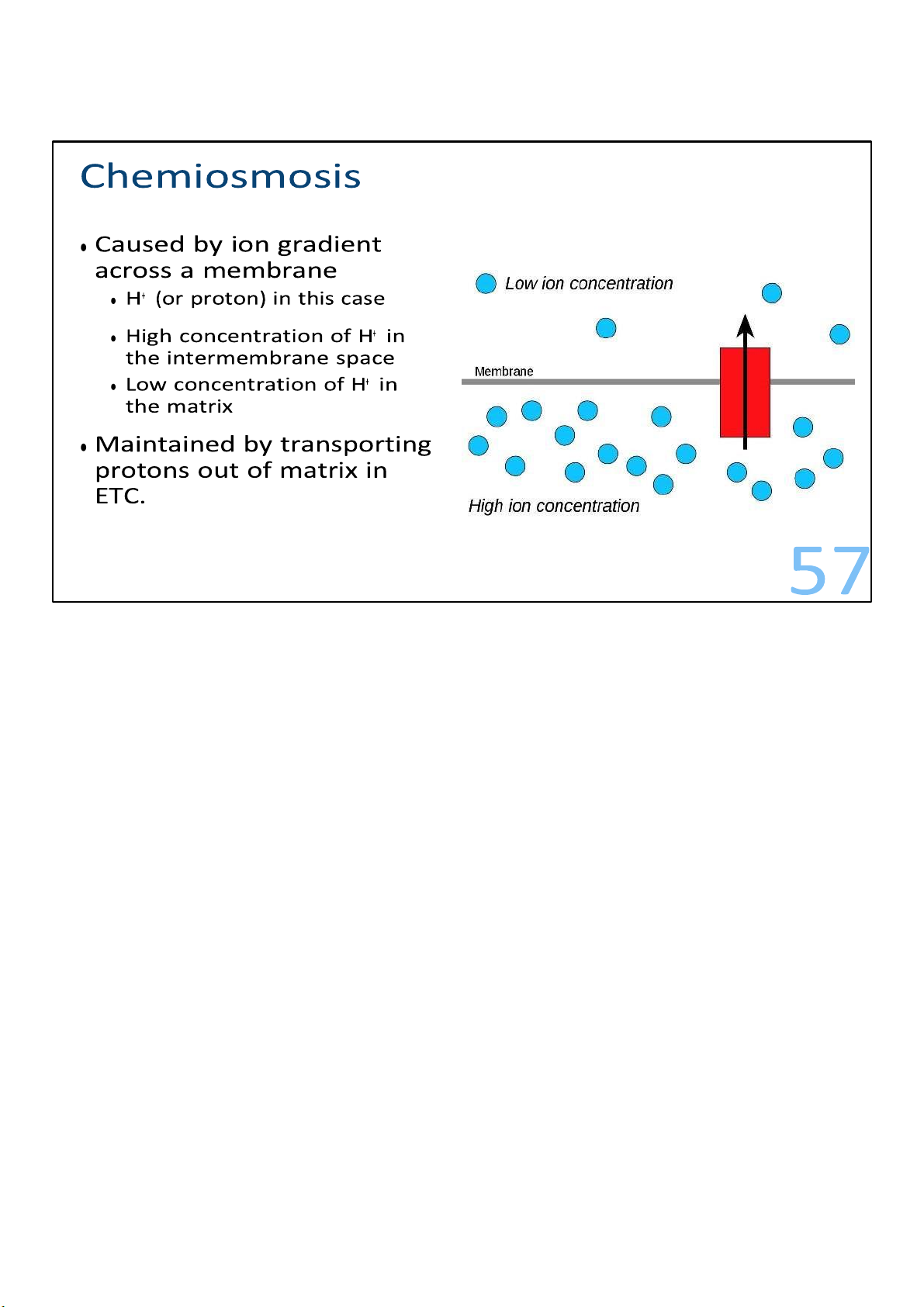
In essence, the ETC is a process where energy released is used to pump
protons out of the mitochondrial matrix and to create a proton
gradient across the inner mitochondrial membrane. It consists
of 4 steps to move the electrons to the final acceptor, usually oxygen. 48 lOMoARcPSD|364 906 32
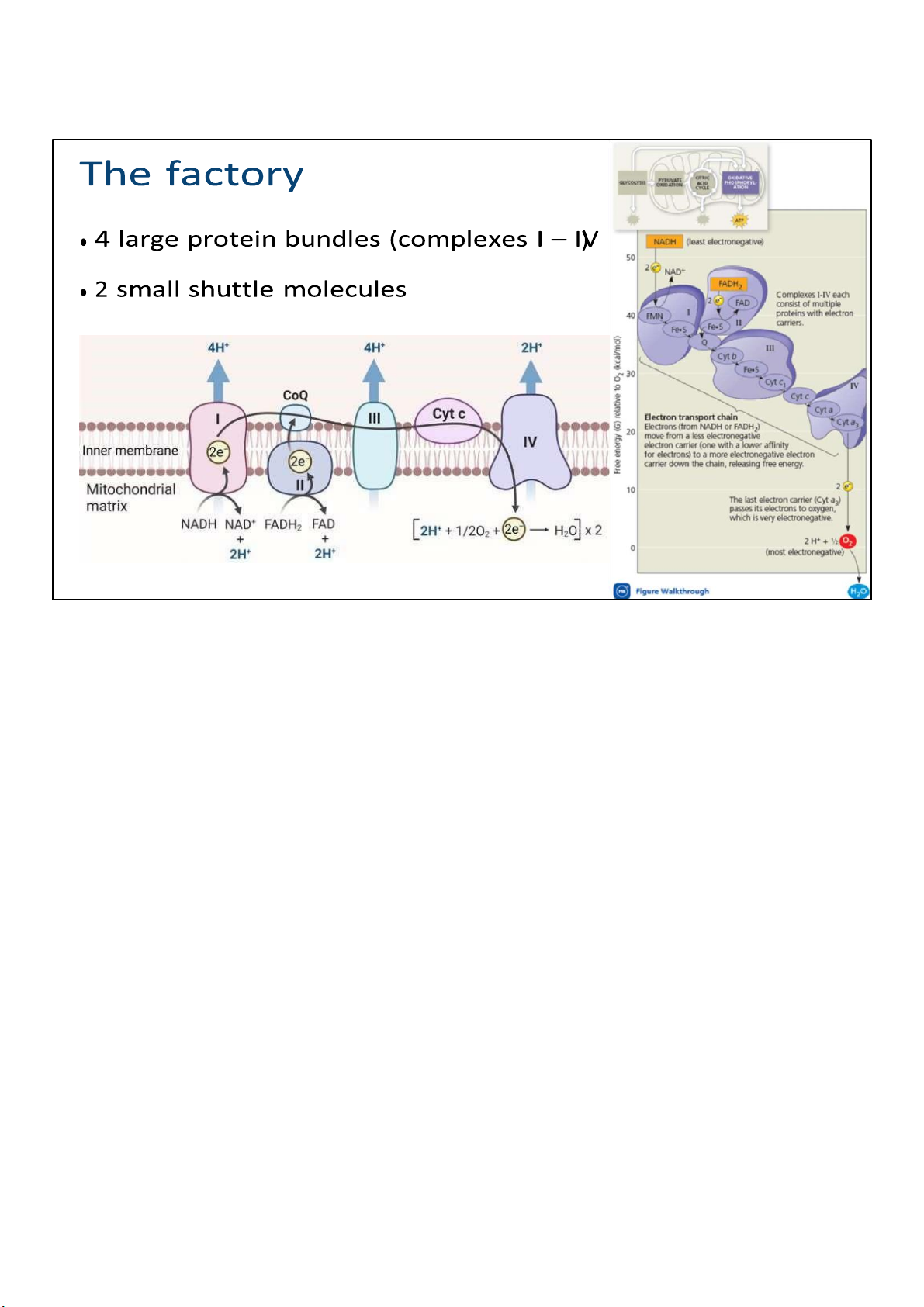
The electron transport chain is a collection of molecules
embedded in the inner membrane of the mitochondrion in
eukaryotic cells. (In prokaryotes, these molecules reside in the
plasma membrane.) The folding of the inner membrane to
form cristae increases its surface area, providing space for
thousands of copies of each component of the electron
transport chain in a mitochondrion.
Most components of the chain are proteins, which exist in
multiprotein complexes numbered I through IV. They’re
considered enzymes, and require cofactors and coenzymes to perform their function. 49 lOMoARcPSD|364 906 32
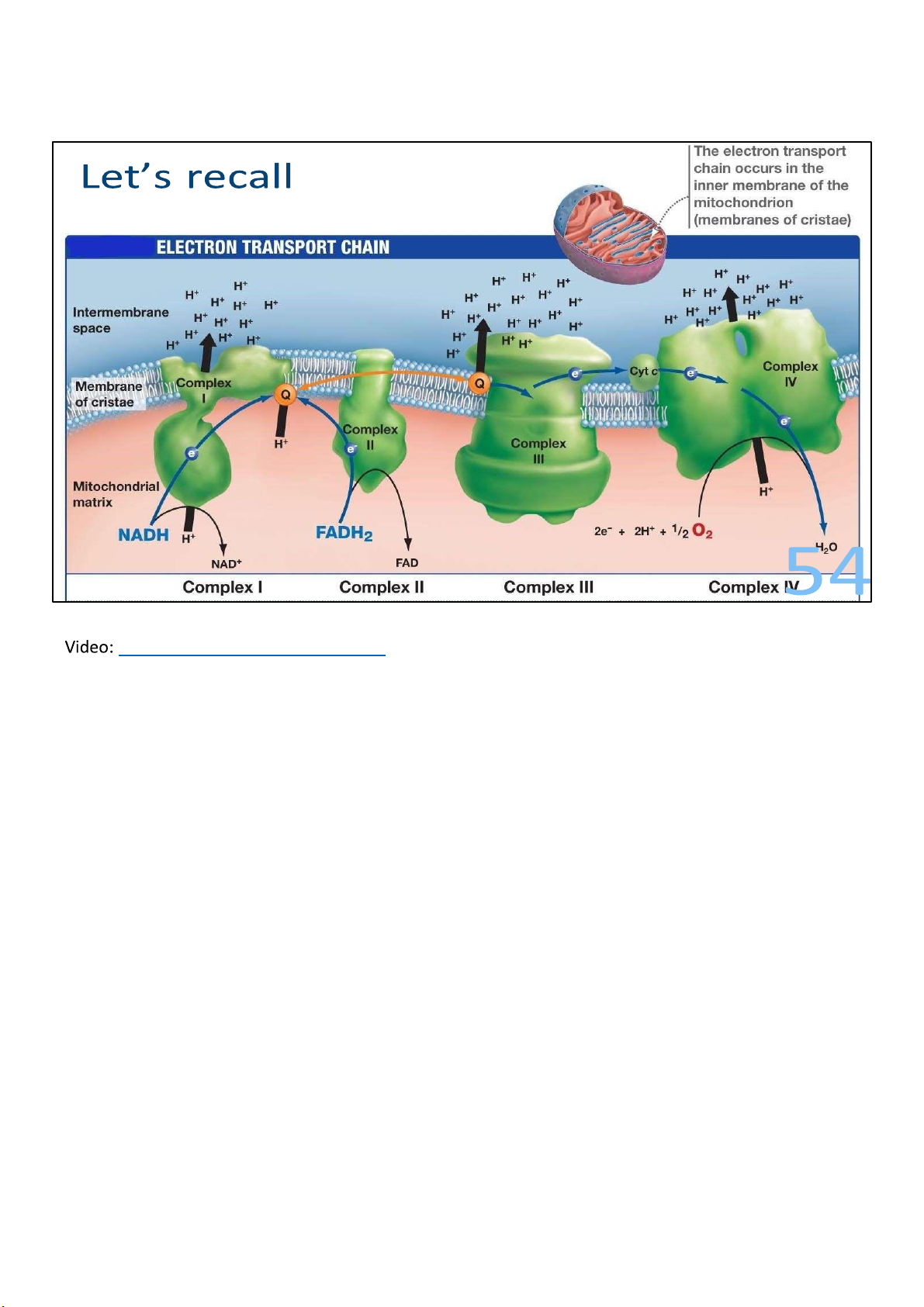
This figure illustrates the way these complexes are embedded
on the inner membrane. The black arrow shows the transport 50 lOMoARcPSD|364 906 32
of electrons, starting from NADH and FADH2, which you
recall are produced in glycolysis and CAC, and ending in
oxygen molecule, which we call the final electron
acceptor. Now, not all ETC ends in oxygen of course, but
for the sake of simplicity and clarity, it’s shown by
default. The blue arrows show the pumping of protons
(H+) out of the matrix and into the intermembrane space.
When I say pumping, it’s implied that energy has to be
spent, right? So where does that energy come from? If
you look at the figure on the right, which plots the
electron carriers in a free energy scale, you can see a
clear extracting of energy after each step. As electrons
travel down the chain to a more electronegative
acceptor, the free energy level drops, which means
some energy is released. Just like how hydroelectricity, a
form of energy, is made when water travels downhill. 49 lOMoARcPSD|364 906 32
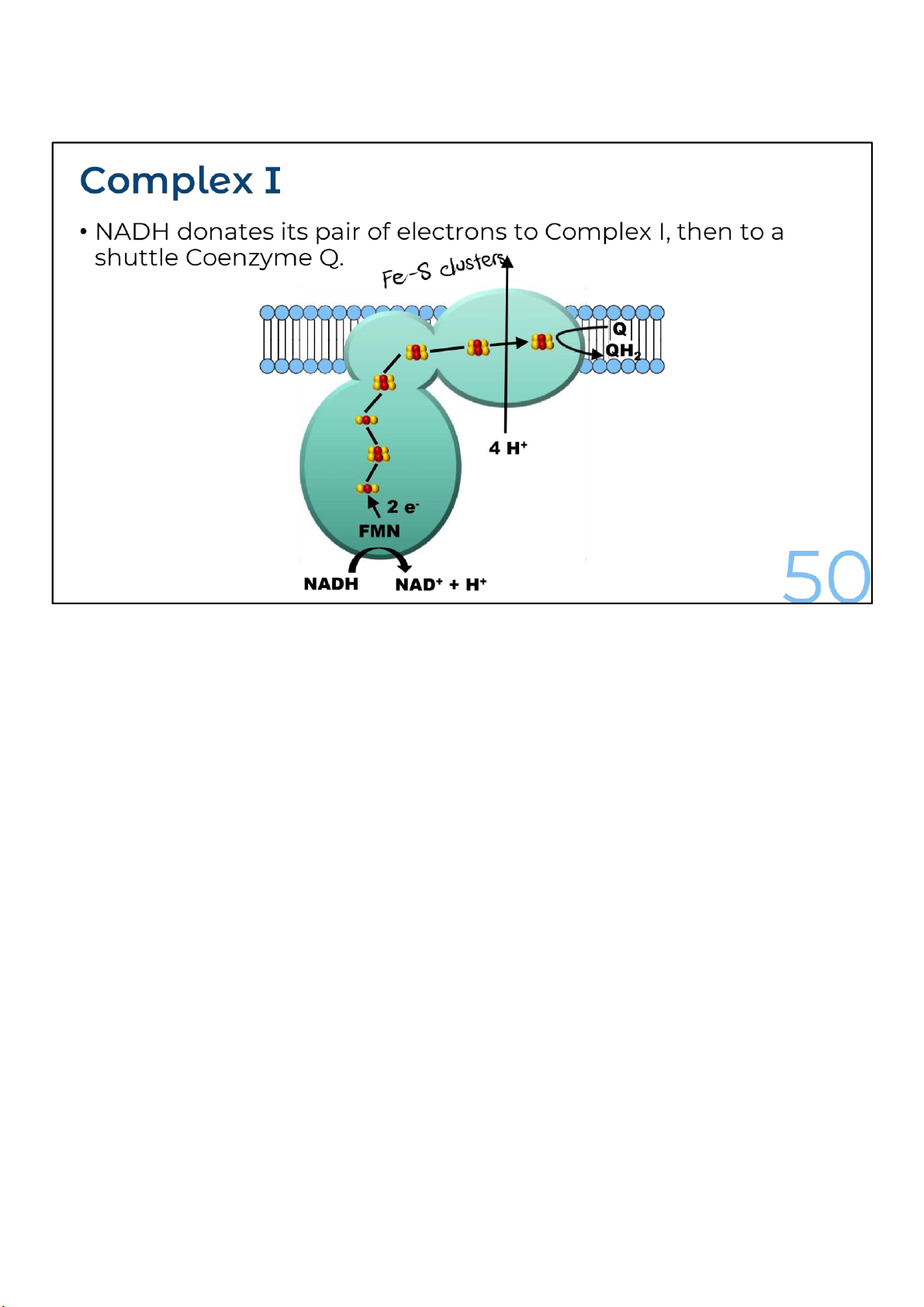
The complex I is a large, multisubunit complex with approx. 40 polypeptide chains. It passes electron
from NADH to coenzyme Q. It contains one flavoprotein with a molecule of flavin mononucleotide
(FMN) and six to seven proteins of iron-sulfur clusters which participate in the process of electron-
transport. During the transport of each pair of the electron from NADH to coenzyme Q, the complex I
pumps four protons across the inner mitochondrial membrane.
Coenzyme Q is a small hydrophobic molecule, the only
member of the electron transport chain that is not a protein.
It’s also known as ubiquinone, and is individually mobile within
the membrane rather than residing in a particular complex,
which is why it’s a form of shuttle. 50 lOMoARcPSD|364 906 32
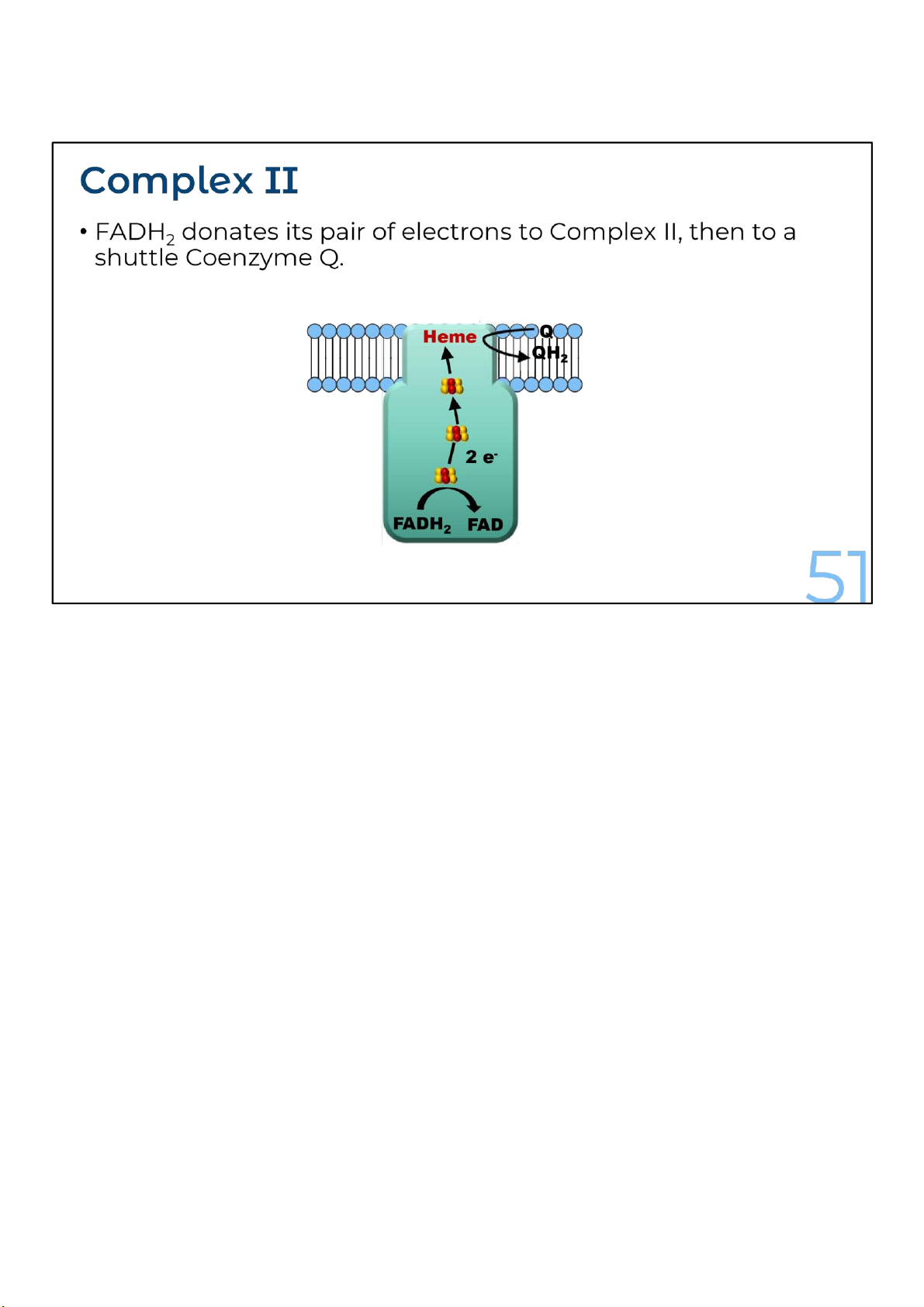
Another source of electrons for the electron transport chain is
FADH2, the other reduced product of the citric acid cycle.
FADH2 adds its electrons from within complex II, at a lower
energy level than NADH does. Consequently, although NADH
and FADH2 each donate an equivalent number of electrons (2)
for oxygen reduction, the electron transport chain provides
about one-third less energy for ATP synthesis when the
electron donor is FADH2 rather than NADH.
Also notice that Complex II does not pump protons during transport of electrons across the inner mitochondrial membrane. 51 lOMoARcPSD|364 906 32
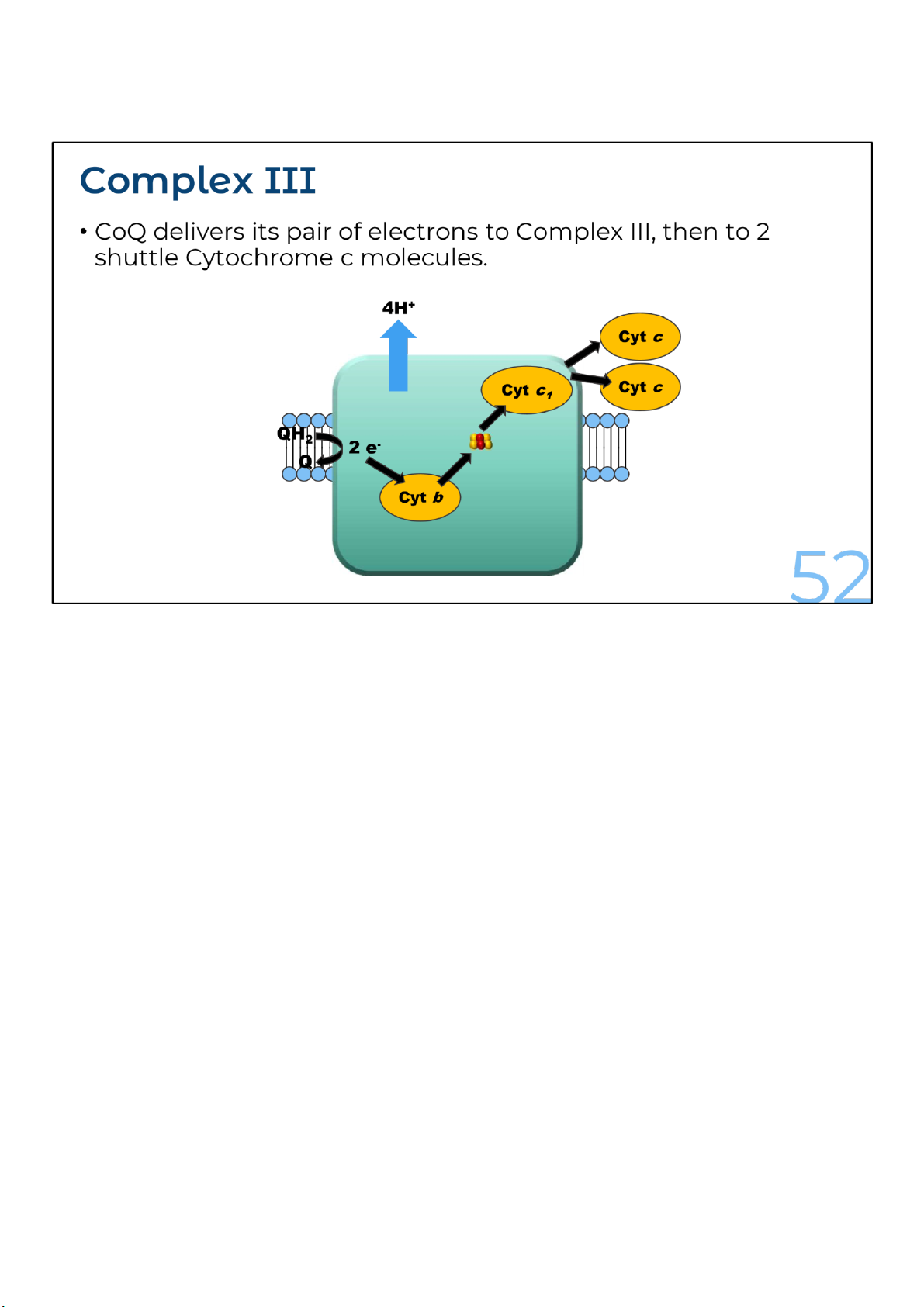
Complex I or complex II donates two electrons to the complex III and regenerates oxidized CoQ. This
process, called the Q cycle, is actually a bit more complicated, but I’ll just show the simplified version with the net results here.
Within complex III, the released electrons are transferred to an iron-sulfur center and then to btype cytochromes or cytochrome c1.
Finally, the two electrons are transferred to two molecules of the oxidized form of cytochrome c. Two
additional protons are translocated from the mitochondrial matrix across the inner mitochondrial
membrane for each pair of electrons transferred.
Cytochromes are heme proteins having distinctive visible-light spectra. The major respiratory
cytochromes are classified as b, c or a depending on the wavelength of the spectral absorption peaks.
Within each class, the cytochromes are distinguished by smaller spectral differences. In the respiratory
electron carriers, there are two b-type cytochromes, cytochrome c and c1 and cytochromes a and a3. 52 lOMoARcPSD|364 906 32
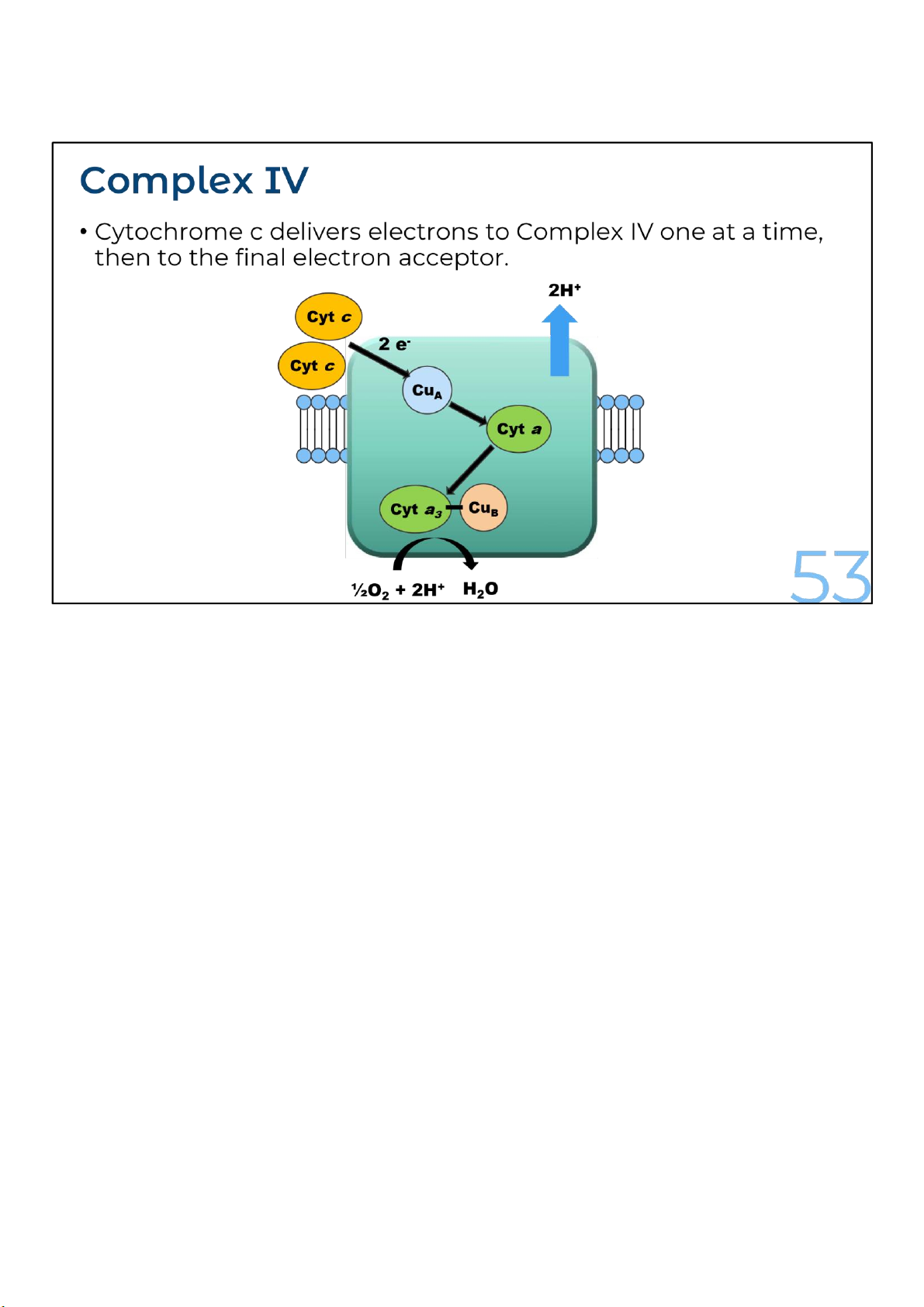
Complex IV or cytochrome c oxidase catalyzes the transfer of electrons from the reduced form of
cytochrome c to molecular oxygen.
Cytochrome c transports electrons, one at a time, to the complex IV. Within this complex, electrons are
transferred, first to a Cua center, then to Cyt a, next to Cub center and Cyt a3 and finally to O2, the
ultimate electron acceptor, yielding H2O. Together, heme a3 and Cub form the active center at which O2 is reduced to H2O.
Two electrons, sequentially released from two molecules of reduced cytochrome c together with two
protons from the matrix, combine with one oxygen atom to form one water molecule. Additionally, for
each electron transferred from cytochrome c to oxygen, one proton is transported from the matrix to
the intermembrane space, or a total of four electrons are transferred for each O2 molecules reduced to two H2O molecules. 53 lOMoARcPSD|364 906 32
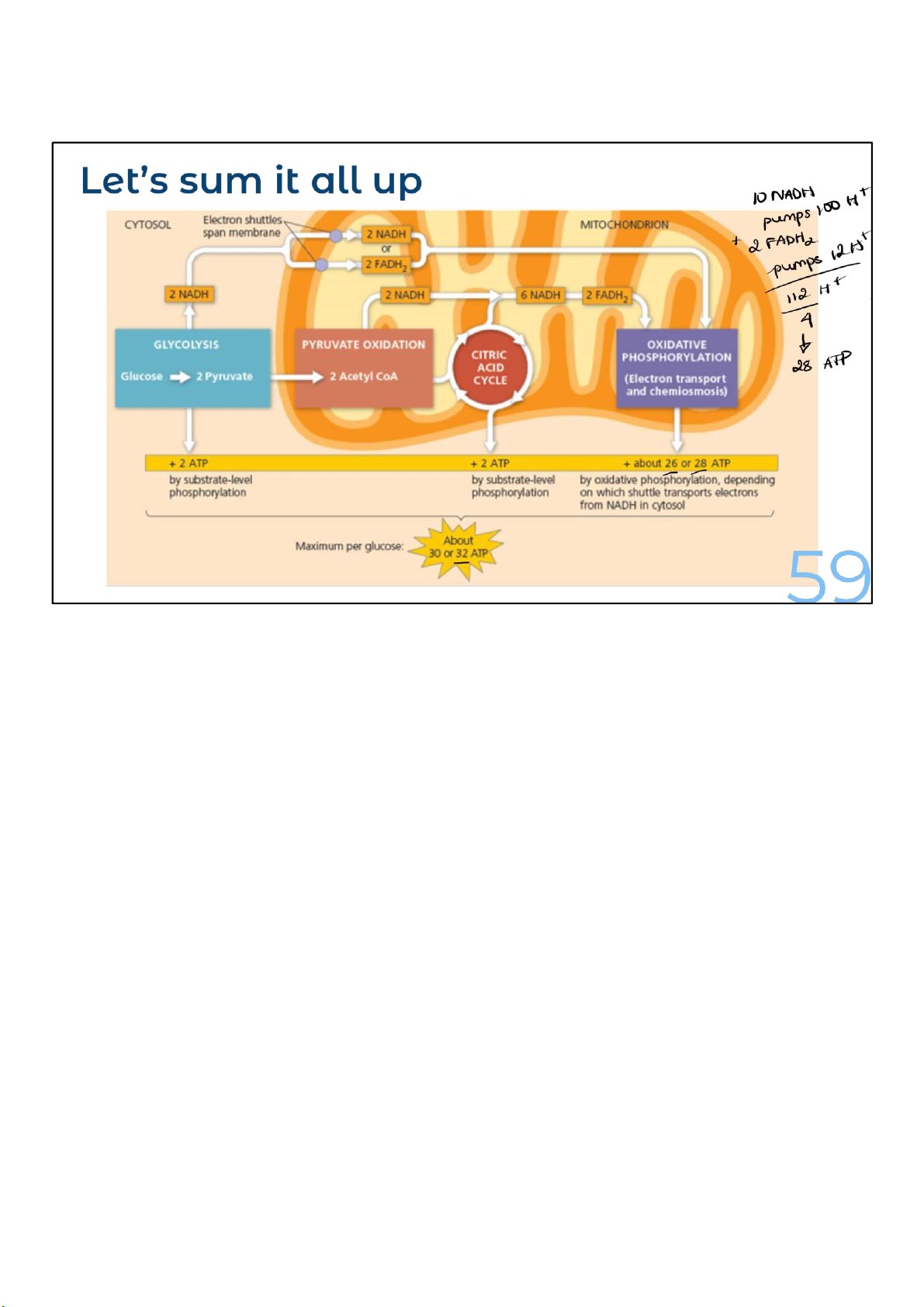
Electron transport chain – YouTube
Starting with glycolysis, moving through the citric acid
cycle, and ending up with oxidative phosphorylation, the
balance sheet for ATP looks like this for each molecule of glucose: -
Glycolysis (glucose to pyruvate): 2 ATP (net) and 2 NADH -
Conversion of 2 pyruvate to 2 acetyl-CoA: 2 NADH - 2
citric acid cycles: 2 GTP (= 2 ATP), 6 NADH, and 2 FADH2
The total so far is 4 ATP, 10 NADH, and 2 FADH2.
Electrons coming from NADH pass through 3 proton-pumping
complexes: I, III, and IV. Electrons entering from FADH2 only
pass through 2 complexes that pump protons, III and IV,
because complex II doesn’t pump any protons. As a result, 54 lOMoARcPSD|364 906 32
electrons from NADH cause more protons to be pumped
across the membrane than electrons from FADH2. In particular, each 55 lOMoARcPSD|364 906 32
NADH oxidized causes 10 protons to be pumped, while
each FADH2 only pumps 6 protons.
So in total, there are 112 protons being pumped after
the ETC finishes with all the NADH and FADH2 from
glycolysis and CAC, if we assume absolute efficiency. 54 lOMoARcPSD|364 906 32 Oxidative phosphorylation 55 55 lOMoARcPSD|364 906 32
The machinery – ATP synthase • Also called Complex V
• Embedded on inner membrane • A pump in “reverse”
• Mushroom-like assembly of 2 multiproteins – stator and rotor
• Proton past through stator stalk to rotor
turn the rotor conformational change in 56 synthesis catalytic knob catalyzing ATP
Compared to electron transport, oxidative phosphorylation is
simple. All of the magic occurs within a remarkable protein
machine called ATP synthase, also known as complex V. ATP
synthase is made up of 2 connected multiprotein assemblies,
which together look like a mushroom. The base of the stalk is
rooted in the mitochondrial inner membrane, while the cap,
or head, projects into the mitochondrial matrix.
As the protons flow back into the matrix through the ATP
synthase, they turn the stalk. This in turn causes
conformational changes in the F1 head, the part of the
enzyme that actually makes the ATP. 56 lOMoARcPSD|364 906 32 57 lOMoARcPSD|364 906 32
ATP synthase in action – YouTube
Around 4 protons are needed to synthesize 1 ATP. 58 lOMoARcPSD|364 906 32 59
Document Outline
- Carbohydrates Metabolism
- Sugars: Glucose & the Carbohydrates
- 10
- Stage 1: Energy investment
- Glycolysis and other pathways
- Stage 1: Energy investment
- 29
- Citric acid cycle
- It’s a cycle
- Citric acid cycle
- 32
Tài liệu liên quan:
-

Genetically Modified Microorganisms for Biofuels | Môn Biochemistry - Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh
123 62 -

Introductory Report on Key Learnings | Môn Biochemistry - Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh
131 66 -

Midterm Review: Enzymes, Metabolism, and Applications | Môn Biochemistry - Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh
119 60 -

Biochemistry Unit Notes: Carbohydrates, Amino Acids, and More | Môn Biochemistry - Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh
125 63 -

Biochemistry Lab Manual for Practical Experiments (Sept 2023) | Môn Biochemistry - Trường Đại học Quốc tế, Đại học Quốc gia Thành phố Hồ Chí Minh
124 62