Chương trình luyện đề thực chiến: Kỹ thuật di truyền và ứng dụng môn Sinh học di truyền | Đại học Đồng Tháp
Kỹ thuật di truyền là thao tác thay đổi gen bằng công nghệ sinh học. Nó bao gồm các phương pháp kỹ thuật dùng để thay đổi nhân tố di truyền của các tế bào, bao gồm sự dịch chuyển gen cùng loài và khác loài để tạo ra những sinh vật mới hoặc hoàn hảo hơn. Tài liệu được sưu tầm gồm 55 trang, giúp các bạn nắm vững kiến thức, rèn luyện kỹ năng và đạt được kết quả tốt trong học tập. Mời các bạn đón xem!
Môn: Sinh học di truyền (DT01) 5 tài liệu
Trường: Trường Đại học Đồng Tháp 374 tài liệu
Tác giả:













Preview text:
Kính mời quý thầy cô và các bạn tham khảo chương trình
LUYỆN ĐỀ THỰC CHIẾN
Có giới hạn người tham gia
CHI TIẾT TẠI: https://forms.gle/xHXk8YHGZuXfRyKL7
CHUYÊN ĐỀ: KỸ THUẬT DI TRUYỀN VÀ ỨNG DỤNG MỤC LỤC
PHẦN 1: MỞ ĐẦU .................................................................................................................. 3
1. Lý do chọn đề tài ................................................................................................................... 3
2. Mục đích của đề tài ................................................................................................................ 4
PHẦN 2: NỘI DUNG .............................................................................................................. 6
1. NHÂN DÒNG ADN ............................................................................................................. 6
1.1. Các enzyme cắt giới hạn ..................................................................................................... 7
1.2. Vector tách dòng ............................................................................................................... 11
1.3. Kỹ thuật nhân dòng ........................................................................................................... 17
1.4. Các ứng dụng của kỹ thuật tách dòng ............................................................................... 23
2. PCR – PHẢN ỨNG CHUỖI TRÙNG HỢP .................................................................... 23
2.1. Các thành phần tham gia vào phản ứng PCR ................................................................... 24
2.2. Phản ứng PCR xảy ra như thế nào .................................................................................... 25
2.3. Các ứng dụng của phản ứng PCR ..................................................................................... 28
3. GIẢI MÃ TRÌNH TỰ ADN .............................................................................................. 29
3.1. Nguyên lý giải mã trình tự ADN ...................................................................................... 29
3.2. Phương pháp hóa học của Maxam-Gilbert ....................................................................... 30
3.3. Phương pháp kết thúc chuỗi trùng hợp nhờ dideoxynucleotide ....................................... 32
3.4. Giải mã trình tự ADN tự động .......................................................................................... 36
4. CÂU HỎI LUYỆN TẬP.....................................................................................................36
PHẦN 3: KẾT LUẬN VÀ ĐỀ NGHỊ....................................................................................52
1. Kết luận.................................................................................................................................52
2. Đề nghị..................................................................................................................................52
TÀI LIỆU THAM KHẢO........................................................................................................53 PHẦN 1: MỞ ĐẦU
1. Lý do chọn đề tài
Kỹ thuật di truyền là thao tác thay đổi gen bằng công nghệ sinh học. Nó bao gồm
các phương pháp kỹ thuật dùng để thay đổi nhân tố di truyền của các tế bào, bao
gồm sự dịch chuyển gen cùng loài và khác loài để tạo ra những sinh vật mới hoặc hoàn hảo hơn.
Những thành tựu rực rỡ của di truyền học đã đem lại những nhận thức mới về cấu
tạo và sự vận hành bộ máy di truyền của cơ thể sống. Cùng với sự phát triển mạnh
mẽ của di truyền học, một lĩnh vực nghiên cứu mới của di truyền học ra đời, đó là
kỹ thuật di truyền hay còn gọi là công nghệ gen hoặc công nghệ di truyền (genetic
engineering). Có thể nói, kỹ thuật di truyền là một tập hợp của nhiều kỹ thuật như
hóa học, sinh học phân tử, vi sinh vật học,... mà trong đó, vai trò hàng đầu thuộc
về các tư duy và phương pháp của di truyền.
Công nghệ di truyền sử dụng các phương pháp sinh học phân tử để tách DNA từ
một cơ thể sống và sau đó cắt, nối các gen trên DNA. Bằng cách như vậy, người
ta có thể loại bỏ các gen không mong muốn và đưa vào các gen mới đặc hiệu theo
chủ ý lựa chọn. Các thao tác cắt, nối trên DNA được thực hiện bên ngoài cơ thể
sống trong các ống nghiệm (in vitro). Phân tử DNA mới được tạo dựng sau các
thao tác cắt, nối có một số đặc điểm khác với phân tử DNA ban đầu được tách ra
từ tế bào sống, được gọi là DNA tái tổ hợp và kỹ thuật này được gọi là kỹ thuật
tái tổ hợp DNA. Sự tái tổ hợp DNA được đánh giá là thành công chỉ sau khi đưa
được phân tử DNA tái tổ hợp vào trong tế bào sống và chúng biểu hiện các hoạt
tính di truyền và ở thế hệ con cháu sẽ mang phân tử DNA tái tổ hợp
Kỹ thuật di truyền đã được áp dụng trong nhiều lĩnh vực khác nhau bao gồm
nghiên cứ, y học, công nghệ sinh học, và nông nghiệp. Trong nghiên cứu, GMO
được dùng để nghiên cứu về chức năng gen qua các phương pháp như làm mất
chức năng, tạo ra chức năng mới, theo dõi biểu hiện gen. Bằng cách loại bỏ một
số gen với chức năng đã được biết đến, chúng ta có thể tạo ra sinh vật mô hình
động vật để nghiên cứu bệnh con người. Ngoài việc có thể tạo ra nội tiết tố, vắc-
xin, và các dược phẩm khác, kỹ thuật di truyền còn có tiềm năng chữa bệnh qua
phương pháp điều trị gen. Những phương pháp kỹ dùng để tạo ra dược phẩm còn
có các ứng dụng công nghiệp khác như sản xuất enzyme để tạo ra thuốc tẩy, phô
mai, và các sản phẩm khác.
Chính vì vậy, kỹ thuật gen thực sự trở thành nhu cầu của sự hiểu biết để con người
có thể sử dụng công nghệ nhằm phục vụ mục đích khác nhau. Ở nước ta, kỹ thuật
gen không chỉ được giảng dạy ở bậc đại học và THPT mà ngày càng thâm nhập
vào nhiều lĩnh vực hoạt động khoa học và lĩnh vực kinh tế khác. Tuy nhiên, ngoài
nội dung kiến thức được dùng để ôn thi đại học thì kỹ thuật gen còn trở thành một
trong những nội dung của các đề thi tuyển chọn học sinh giỏi Quốc gia và Olympic
Sinh học. Đây là nội dung tương đối khó, học sinh phải sử dụng nhiều kiến thức
chuyên ngành và kiến thức thực tiễn để giải quyết vấn đề này, một nội dung ít
được tiếp cận trong chương trình sinh học THPT.
Với mong muốn nâng cao chất lượng của HSG, cũng như cung cấp cho giáo viên
và học sinh nguồn tư liệu để tham khảo, giảng dạy, học tập và quan trọng hơn là
hoàn thiện kiến thức của bản thân nên Tôi đã chọn nội dung “ Nguyên lý kỹ thuật
di truyền và Ứng dụng” làm đề tài nghiên cứu.
2. Mục đích của đề tài -
Cung cấp nguồn tư liệu tham khảo cho giáo viên và học sinh chuyên
tronggiảng dạy và bồi dưỡng học sinh giỏi:
+ Giúp giáo viên và học sinh có cái nhìn tổng quan và chuyên sâu về phần kỹ thuật di truyền.
+ Hiểu sâu hơn về các nguyên lý của kỹ thuật di truyền cơ bản và ứng dụng
trong việc phân tích gen và các sản phẩm của gen. -
Giúp học sinh chuyên biết cách đọc tài liệu để phát huy năng lực theo hướng
tựhọc, tự nghiên cứu: khái quát vấn đề, tự đặt các câu hỏi, hiểu được nguyên lý
của các phương pháp và khả năng ứng dụng trong thực tế. -
Xây dựng hệ thống câu hỏi liên quan đến các kỹ thuật di truyền, từ đó rèn
luyện kĩ năng trả lời câu hỏi cho học sinh. -
Xây dựng được mạch kiến thức cơ bản của chuyên đề, có thể áp dụng để
dạynền cho mọi đối tượng học sinh. PHẦN 2: NỘI DUNG 1. NHÂN DÒNG ADN
Nhân dòng gen là khái niệm chỉ một nhóm các phương pháp được sử dụng
để phân lập một một trình tự gen (ADN) đặc hiệu từ hỗn hợp các phân tử ADN
ban đầu được tách chiết từ các mẫu sinh học vốn có cấu trúc phức tạp, kích thước
lớn; khuếch đại (sao chép) trình tự lên một số lượng lớn đủ để có thể tiến hành
phân tích về cấu trúc và chức năng của gen tương ứng.
Khả năng tinh sạch các đoạn gen (ADN) đặc hiệu có số lượng đủ lớn là cần
thiết để có thể “thao tác” các đoạn gen đó vì các mục tiêu nghiên cứu khác nhau.
Chẳng hạn, từ các phân đoạn ADN được tách dòng, người ta có thể tạo ra các phân
tử ADN tái tổ hợp mang các phân đoạn ADN mang nguồn gốc khác nhau. Các
phân tử ADN tái tổ hợp mới có thể làm thay đổi mức độ biểu hiện bình thường
của một gen (chẳng hạn bằng việc dung hợp giữa một trình tự mã hóa của một loài
này với trình tự promoter của một loài khác) hoặc thậm chí mã hóa tổng hợp một
loại protein “dung hợp” mới (protein lai) mang các trình tự axit amin từ các protein
có nguồn gốc khác nhau. Hiện nay, các kỹ thuật tách dòng phân tử (bao gồm cả
PCR) đã trở thành các công cụ thiết yếu trong nghiên cứu về sự điều hòa và biểu
hiện của các gen và hệ gen ở các loài sinh vật khác nhau.
Quá trình tách dòng ADN và tạo nên các phân tử ADN tái tổ hợp điển hình
thường liên quan đến việc sử dụng các véctơ là trình tự mang thông tin điều khiển
hoạt động nhân lên (khuếch đại) và/hoặc biểu hiện trong tế bào của phân tử ADN
tái tổ hợp mang đoạn ADN cài (đoạn trình tự được phân lập) bao gồm trình tự gen
được quan tâm nghiên cứu. Các “công cụ” chính để tạo nên các phân tử ADN tái
tổ hợp là các enzym giới hạn giúp cắt các phân tử ADN tại các vị trí xác định và
các enzym nối cho phép ghép nối các phân đoạn ADN có nguồn gốc khác nhau
với nhau. Bằng việc tạo nên các phân tử ADN tái tổ hợp có thể tự nhận lên trong
tế bào chủ, một đoạn ADN cài xác định nào đó có thể được phân lập, tinh sạch và
nhân lên thành một số lượng lớn các bản sao.
Tiếp theo đây, chúng ta sẽ mô tả bằng cách nào các phân tử ADN được cắt,
tái tổ hợp và nhân lên, đồng thời đề cập đến việc xây dựng thư viện hệ gen gồm
tập hợp các phân tử ADN lai chứa các đoạn cài xuất phát từ một hệ gen cần được
thiết lập để phục vụ cho việc nghiên cứu, phân tích một hệ gen. Thông thường một
thư viện hệ gen được thiết lập bằng việc sử dụng chung cùng một loại véctơ mang
các phân đoạn ADN cài khác nhau. Chúng ta sẽ thấy bằng cách nào các phân đoạn
ADN đặc hiệu có thể được xác định và phân lập từ các thư viện hệ gen.
1.1. Các enzyme cắt giới hạn
1.1.1. Enzyme giới hạn
Là một enzyme endonuclease có vị trí nhận biết điểm cắt ADN đặc hiệu.
Những enzyme này phân huỷ liên kết phosphodieste của bộ khung ADN mạch đôi
mà không gây tổn hại đến bases. Các liên kết hóa học mà bị enzyme này cắt có
thể được nối trở lại bằng loại enzyme khác là các ligases, vì thế các phân đoạn
giới hạn (sản phẩm của phản ứng cắt RE) mà bị cắt từ các nhiễm sắc thể hoặc gene
khác nhau có thể được ghép cùng nhau nếu có trình tự đầu dính bổ sung với nhau.
Nhiều kỹ thuật sinh học phân tử và kỹ thuật di truyền đều dựa vào các enzyme
giới hạn. Phân loại enzyme giới hạn
Các nhà sinh hóa chia enzyme cắt giới hạn nói chung thành ba loại, gọi là
Loại I, Loại II và Loại III. Đối với hai loại I và III, cả hoạt tính phân giải acid
nucleic hay phân giải nhóm methyl đều thực hiện chung bởi một phức hợp enzyme
lớn. Mặc dù những enzyme thuộc hai loại này cũng nhận biết những trình tự ADN
đặc hiệu, vị trí cắt thường cách xa vị trí nhận biết, có khi đến cả trăm base. Chúng
cũng cần ATP để hoạt động. Những enzyme này bắt đầu bằng việc kiểm tra tình
trạng methyl hóa của 2 adenine trong vùng nhận biết. Nếu cả hai adenine đều
không được methyl hóa (dấu hiện cho thấy đây là ADN ngoại lai), phức hợp
enzyme thay đổi cấu hình và thực hiện hoạt tính phân giải. Tuy nhiên, nếu một
trong hai adenine được methyl hóa, chứng tỏ là ADN của tế bào, enzyme khi đó
sẽ thực hiện chức năng của một enzyme methyl hóa cho gốc adenine còn lại để
duy trì sự ổn định cho ADN bộ gene. Với enzyme giới hạn loại II, chức năng phân
giải của nó không liên quan đến chức năng methyl hóa hay phân giải nhóm methyl,
và vị trí cắt cũng nằm ngay bên trong hay kế cạnh vị trí nhận biết. Ngày nay người
ta biết rất nhiều enzyme khác nhau loại này và chúng là một trong những công cụ
sinh học phân tử thiết yếu, đặc biệt thường gặp trong các ứng dụng dòng hóa gene hay phân tích ADN.
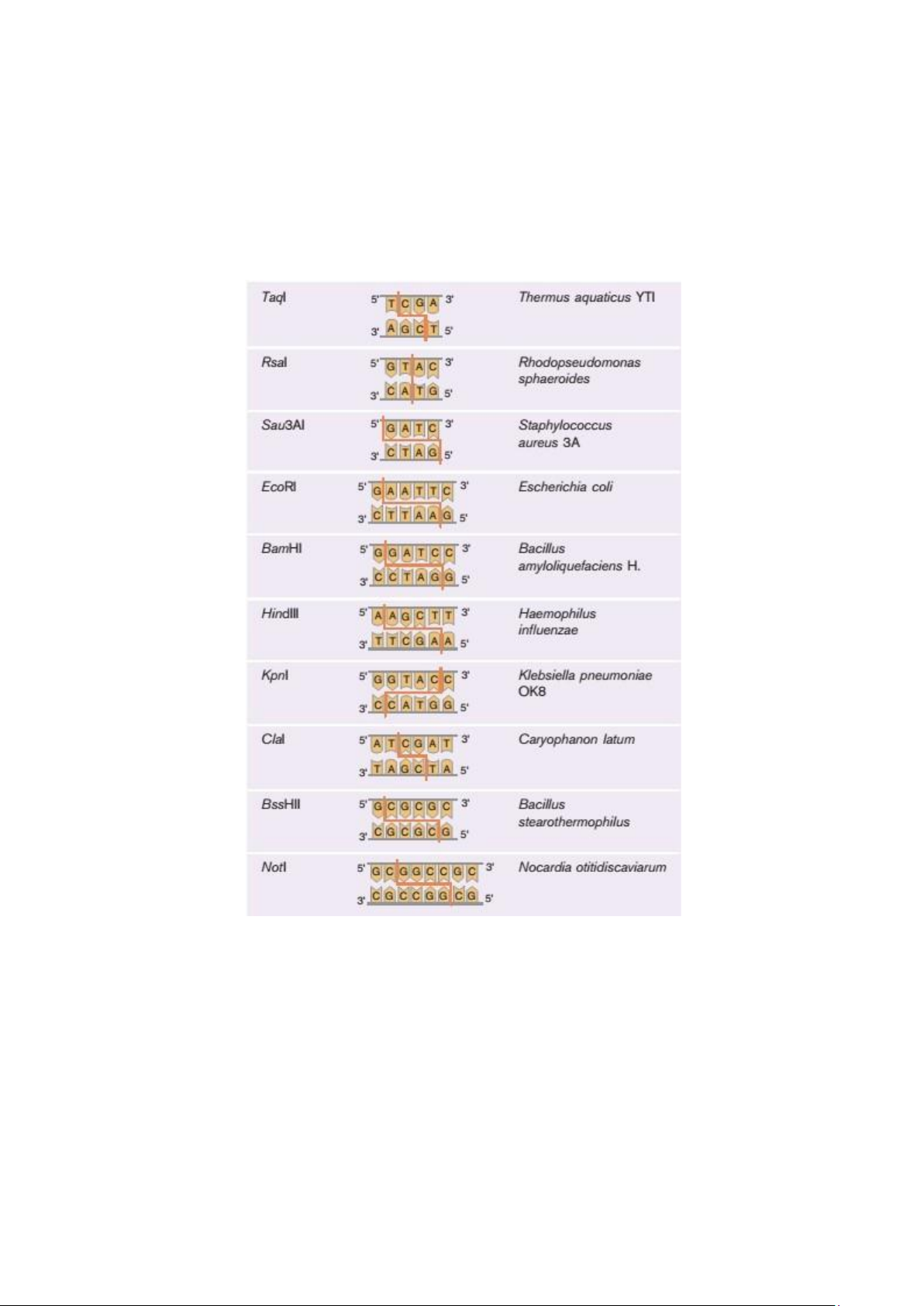
Một số enzyme cắt giới hạn (RE)
Vị trí điểm cắt
Enzyme giới hạn chỉ cắt các trình tự lặp đối xứng khi đọc theo chiều 5´-3´
trên mạch ADN (palindrome) gọi là trình tự nhận biết. Vị trí điểm cắt của enzyme
giới hạn có thể nằm trong hoặc ngoài trình tự nhận biết này. Một số enzyme tạo ra
các vết cắt trên mạch đối diện tức thời, tạo ra các đoạn ADN "đầu bằng". Hầu hết
các en đều tạo ra các vết cắt hơi chéo nhau (hình chữ chi), tạo ra các "đầu dính".
Các enzyme giới hạn có ba chức năng quan trọng, mỗi chức năng cắt ADN bằng các cơ chế khác nhau.
(a) Đầu bằng (Rsal)
(b) Đầu dính 5’ (EcoRI)
(c) Đầu dính 3’ (Kpnl)
2.1.2. Các loại enzyme cắt giới hạn tạo các đoạn ADN khác nhau
Điện di trên gel (gel electrophoresis) áp dụng trong sinh học phân tử là một
kĩ thuật để phân tích các phân tử ADN, RNA hay protein dựa trên các đặc điểm
vật lý của chúng như kích thước, hình dạng hay điểm đẳng điện tích (isoelectric
point). Kĩ thuật này sử dụng một dung dịch đệm (buffer) để dẫn diện và tạo điện
trường đều, một bản gel (thường là agarose hay polyacrylamide) đóng vai trò là
thể nền để phân tách các phân tử, và các chất nhuộm khác nhau (ethidium bromide,
bạc, xanh Coomassie) để phát hiện vị trí các phân tử trên gel sau khi điện di.
Kĩ thuật điện di hoạt động nhờ vào lực kéo của điện trường tác động vào
các phân tử tích điện và kích thước lỗ của thể nền (gel). Gel cấu tạo bởi các chuỗi
cao phân tử (polymer) được liên kết chéo với nhau tạo thành một hệ thống mạng
lưới với kích thước các mắc lưới tùy thuộc vào nồng độ chất cao phân tử (agarose,
polyacrylamide) và phản ứng tạo liên kết chéo. Các phân tử được phân tách khi di
chuyển trong gel với vận tốc khác nhau nhờ vào sự khác nhau của (a) lực của điện
trường tác động lên chúng (nếu các phân tử tích điện khác nhau) (b) kích thước
của phân tử so với kích thước lỗ của gel và (c) hình dạng, độ cồng kềnh của phân tử.
Gel agarose: Là một polysaccharide, thường được chiết xuất từ rong biển đỏ nhất
định. Nó là một polymer tuyến tính được tạo thành từ đơn vị lặp đi lặp lại của
agarobiose, là một disaccharide tạo thành từ D -galactose và 3,6-anhydro- L -
galactopyranose. Agarose là một trong hai thành phần chính của thạch, và được
tinh chế từ thạch bằng cách loại bỏ thành phần khác của agar, agaropectin. Agarose
có cấu trúc dạng lưới ba chiều các kênh có đường kính từ 50 nm đến> 200 nm tùy
thuộc vào nồng độ agarose được sử dụng - nồng độ cao hơn mang lại đường kính
lỗ chân lông trung bình thấp hơn. Cấu trúc 3-D được tổ chức cùng với liên kết
hydro và do đó có thể bị gián đoạn do làm nóng trở lại trạng thái lỏng.
Quy trình điện di gel trên agarose
Chuẩn bị dung dịch đệm Agarose được sử dụng làm Các lỗ trên bản gel là các và bản gel
chất nền để phân tách các giếng chứa ADN đoạn ADN
Nạp mẫu vào các giếng
ADN tích điện âm di chuyển
Nhuộm bản gel với
trên bản gel về phía cực dương ethyldium-bromid
Vì có cấu trúc dạng lưới phù hợp nên agarose thường được sử dụng trong sinh học
phân tử để tách các phân tử lớn, đặc biệt là ADN, bằng điện di. Các tấm gel agarose
(thường là 0,7 - 2%) đối với điện di được chuẩn bị dễ dàng bằng cách đổ dung
dịch ấm, lỏng vào khuôn. Một loạt các agaroses khác nhau của trọng lượng phân
tử khác nhau và tài sản là thương mại có sẵn cho mục đích này. Agarose cũng có
thể được hình thành thành các hạt và được sử dụng trong một số phương pháp sắc
ký để làm sạch protein.
Bản gel sau khi nhuộm, rửa nước thì được đặt vào máy
có tia soi UV để quan sát kết quả điện di.
Các phân tử ADN có kích thước càng lớn (khối lượng
phân tử lớn) thì tốc độ dịch chuyển càng chậm.
Đoạn ADN mang kích thước nhất định sẽ dịch chuyển
ở các tốc độ khác nhau qua các bản gel chứa các nồng độ agarose khác nhau
Agarose có khả năng phân tách các phân tử có kích thước
lớn khác nhau trong khoảng 20kb.
Mời các bạn cùng theo dõi những bước cơ bản trong
quá trình điện di ADN trên gel agarose qua link video sau
https://www.youtube.com/watch?v=BKgi6aJMIWw
1.2. Vector tách dòng
Vector tách dòng (cloning vector) là một phân tử ADN có kích thước nhỏ cho
phép cài gắn một đoạn ADN ngoại lai vào nhằm mục đích nhân đoạn ADN ngoại
lai lên với số lượng lớn.
Các đặc điểm cần có của một véctơ tách dòng: -
Có khả năng cài gắn các đoạn ADN ngoại lai -
Có kích thước nhỏ để xâm nhập tế bào chủ -
Có thể tái bản độc lập không phụ thuộc vào bộ gen của tế bào chủ. -
Có chứa các gen chỉ thị, chọn lọc (gen kháng kháng sinh, gen chỉ thị màu...)
Tùy thuộc vào kích thước đoạn ADN mà chúng ta muốn cài mà sử dụng các
vector tách dòng khác nhau. Các loại vector tách dòng thường dùng như là
Plasmid, Cosmid, Phage, BAC (Bacterium artificial chromosomes), YAC (yeast artificial chromosomes) Vector Tế bào chủ Đoạn ADN cài Lamda phage E.coli 5 – 25 kb Lamda cosmids E.coli 35 – 45 kb P1 phage E.coli 70 – 100 kb PACs E.coli 100 – 300 kb BACs E.coli < 300 kb YACs Saccharomyces 200 – 2000 kb cerevisae
1.2.1. Vector chuyển gen là plasmid
Các plasmid là những mẫu ADN nhỏ, ngắn, dạng vòng (khép kín), sợi đôi
nằm ngoài nhiễm sắc thể, được tìm thấy đầu tiên trong tế bào một số vi khuẩn.
Chúng sao chép được là nhờ một số enzyme có mặt trong tế bào vi khuẩn và không
phụ thuộc vào sự sao chép nhiễm sắc thể vi khuẩn. Tùy các kiểu của plasmid mà
số bản sao plasmid bởi vi khuẩn sẽ khác nhau. Một số plasmid chỉ có một bản sao
duy nhất vì chúng tự tái bản chỉ một lần trong mổi lần phân bào.
Plasmid pGEX-3x là một vector tách dòng phổ biến.
Một số khác có số bản sao lớn vì chúng tái bản được nhiều lần trong mỗi
chu kỳ phân bào. Những plasmid có từ 10 đến 100 bản sao trong tế bào chủ được
xem như plasmid có bản sao cao. Plasmid khác có từ 1 đến 4 bản sao trong tế bào
chủ, được xếp vào nhóm có bản sao thấp. Trong sinh vật eucaryote, plasmid chỉ
có trong tế bào nấm men.
Mỗi plasmid đều có một chuỗi mã di truyền (sequence) mang chức măng
tự tái bản ADN. Nếu không có vị trí khởi đầu phiên mã (ori) này, ADN không thể
tự tái lập trong tế bào chủ. Với tính chất tự tái bản, plasmid là một vector chuyển
gen để nhân dòng ADN cần thiết.
Do kích thước nhỏ nên plasmid chỉ chứa rất ít gen chọn lọc, thường đặc tính
chọn lọc là kháng kháng sinh. Từ khi được phát hiện đến nay, các plasmid không
ngừng được cải tiến và ngày càng được có thêm nhiều đặc tính quí cho việc tạo dòng.
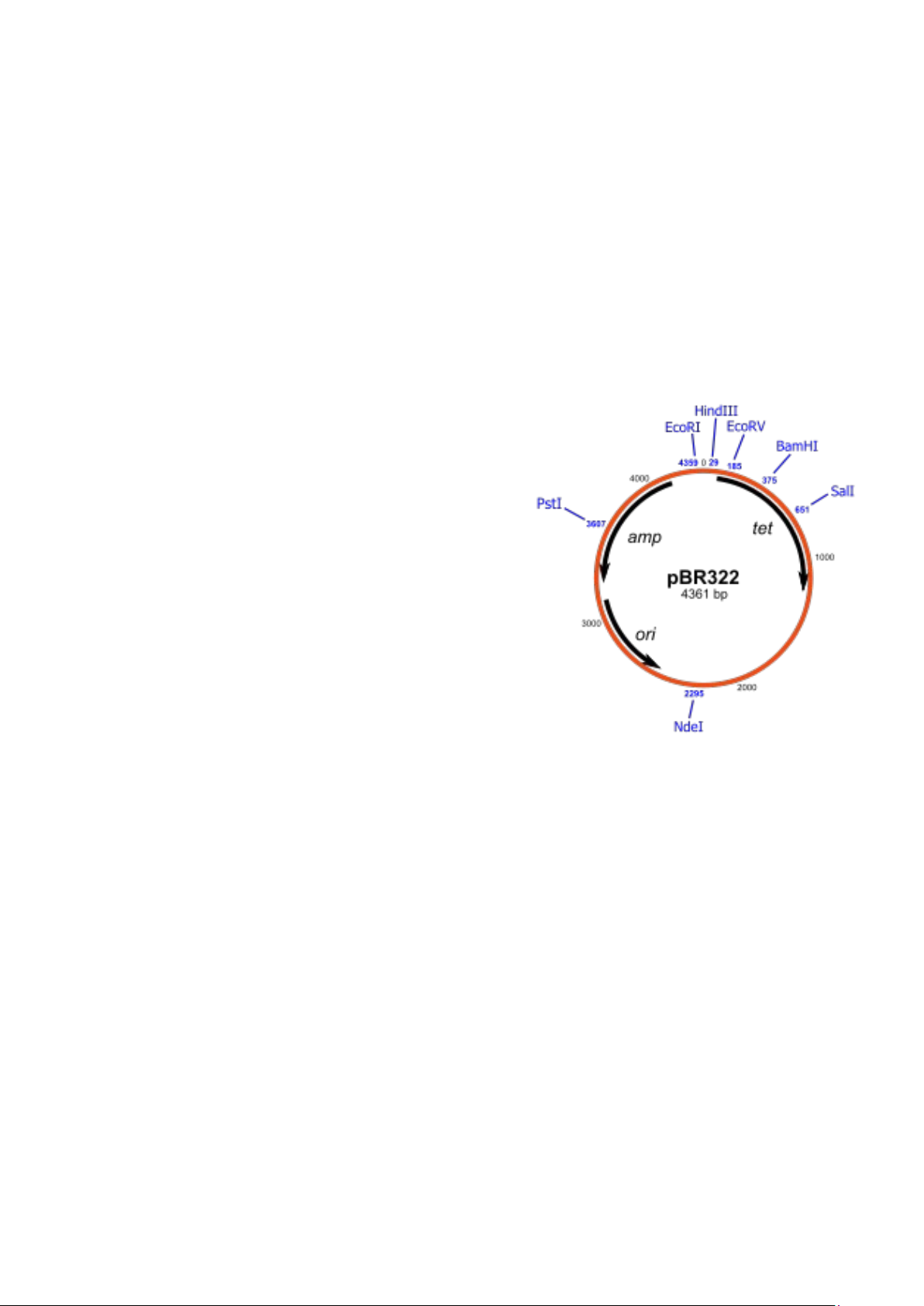
Trong phòng thí nghiệm, người ta sử dụng các plasmid nhân tạo mà được tạo ra
từ các plasmid tự nhiên và cài thêm một số chuỗi ADN.
pBR322 có kích thước 4.363bp và hai gen
kháng thuốc, một chống chịu được ampicilline
(ampR) và một chống chịu được tetracycline (tetR).
Có nhiều điểm nhận biết bởi enzyme cắt hạn
chế và trong số đó có nhiều điểm nhận biết
nằm trong gen kháng kháng sinh. Ví dụ, gen
kháng ampicilline có ba trình tự nhận biết bởi
ba enzyme cắt hạn chế là PstI, PvuI, ScaI. Còn
gen kháng tetracycline có sáu điểm nhận
biết là: EcoRV, BamHI, SphI, SlaI, XmaIII, Cấu tạo plasmid pBR322 NnuI.
Việc cài ADN lạ được tiến hành một trong hai gen kháng thuốc. Nếu chỉ còn một
gen kháng với kháng sinh là do một trong hai gen đã không nhận đoạn cài. Điều
này cho phép để chọn lựa các plasmid tái tổ hợp. Một ứng dụng nữa của pBR322
là chúng có số bản sao lớn. Thực nghiệm cho thấy cứ 15 plasmid tái tổ hợp được
biến nạp trong E. Coli, số lượng này có thể tăng lên từ 1.000 đến
3.000 bản sao trong điều kiện nuôi cấy tốt. 1.2.2.
Các vector chuyển gen là phage
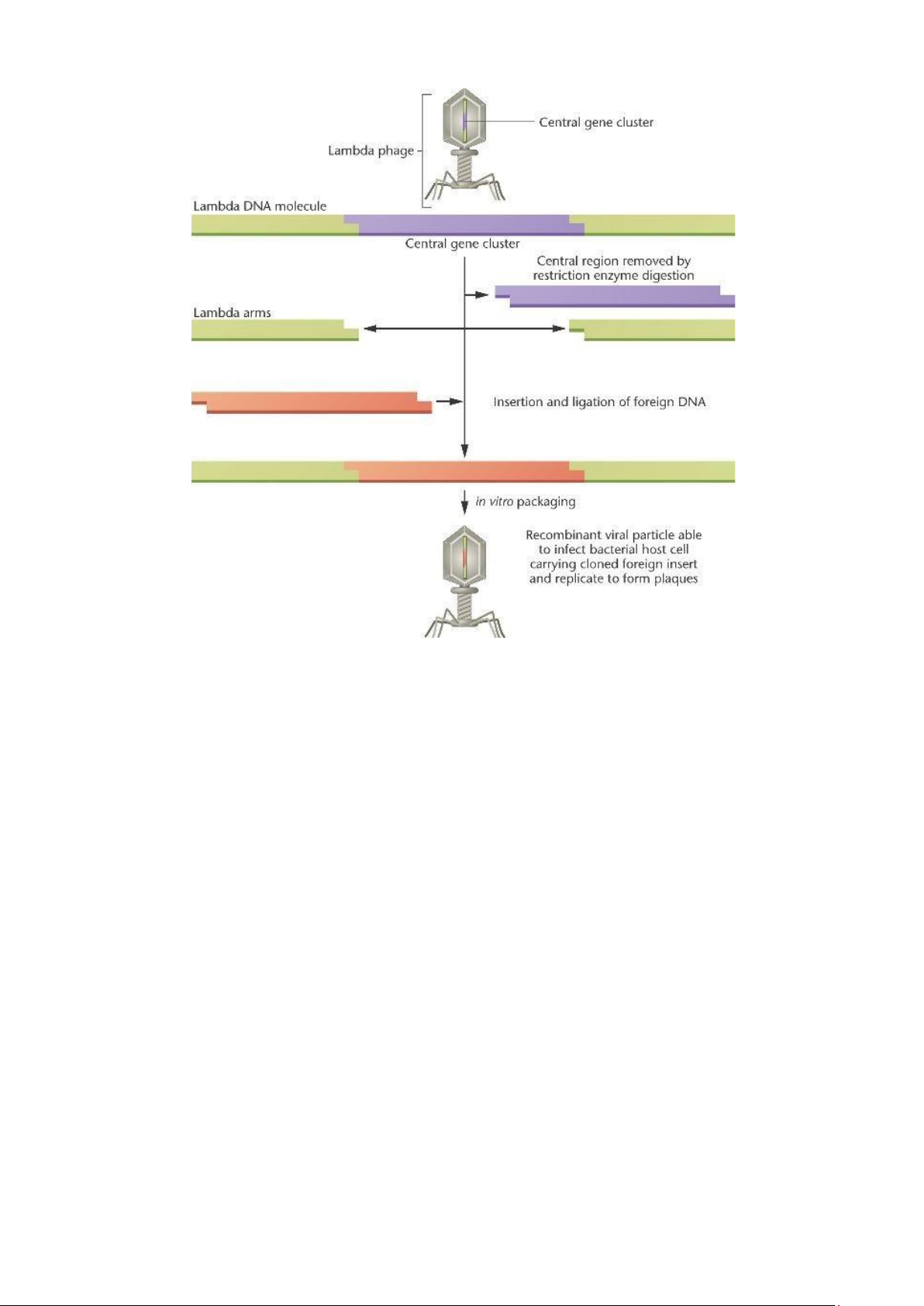
Lamda phage làm vector chuyển những đoạn gen có kích thước nhỏ
Phage (thực khuẩn thể) là virus xâm nhiễm vi khuẩn làm phân giải vi khuẩn.
Việc sử dụng phage làm vector chuyển gen có nhiều ưu điểm hơn so với vector là plasmid
- Dễ xâm nhập vào vi khuẩn,
- Khả năng nhân lên nhanh trong tế bào chủ,
- Khả năng tiếp nhận đoạn ADN lạ lớn hơn plasmid.
Tuy nhiên, việc sử dụng phage có nhiều bất lợi như
- Thao tác ghép ADN lạ phức tạp,
- ADN tái tổ hợp không tạo thành khuẩn lạc như ADN tái tổ hợp là plasmid,
màthành đĩa phân giải xuất hiện trên mặt thạch phủ đầy vi khuẩn
Phần lớn các nhóm phage sử dụng làm vector đều bắt nguồn từ phage λ thuộc thế hệ thứ nhất.
1.2.3. Các vector chuyển gen khác
- Các cosmid: là những vector lai nhân tạo từ một plasmid với các trình tự cos
của phage λ (được sử dụng từ năm 1978). Các trình tự cos này điều khiển sự đóng
gói ADN tái tổ hợp vào đầu của phage. Khi bao gói các vùng cos đều bị cắt, chỉ
còn một phần ADN của cosmid được giới hạn bởi các đầu dính với đoạn cài ADN
lạ được bao gói. Trong phản ứng bao gói in vitro, các protein cần thiết cho sự tạo
thành đầu và đuôi phải được thêm vào để cho các phage có thể tự hợp thành.
Vector chuyển là cosmid
Cosmid là những plasmid có các vùng giới hạn mà tại đây, người ta có thể
cài lắp ADN lạ và một gen chống chịu ampicilline. Kích thước cosmid ≈ 5kb do
đó, nó có thể nhận được đoạn cài 35÷45kb . Vào thời điểm gây nhiễm E. Coli,
ADN tái tổ hợp được phóng vào vi khuẩn. Trong vi khuẩn các đầu dính sẽ bắt cặp
tạo ra cosmid tái tổ hợp khép kín dạng vòng và tái bản như một plasmid. Ưu điểm
và ứng dụng của cosmid: Cũng như phage, cosmid cho khả năng xâm nhiễm tế
bào vi khuẩn lớn, nhận đoạn cài có kích thước lớn. Trong tế bào chủ, nó tự nhân
bản như plasmid. Cho nên người ta nhận được những khuẩn lạc, chứ không phải
đĩa phân giải trên mặt thạch, thuận lợi cho việc quan sát. Với những tiện lợi trên,
người ta dùng cosmid để tách dòng từ những gen lớn để tạo ngân hàng genom (bộ
gen). Những vi khuẩn mang vector này có khả năng chống chịu với môi trường có ampiciline.
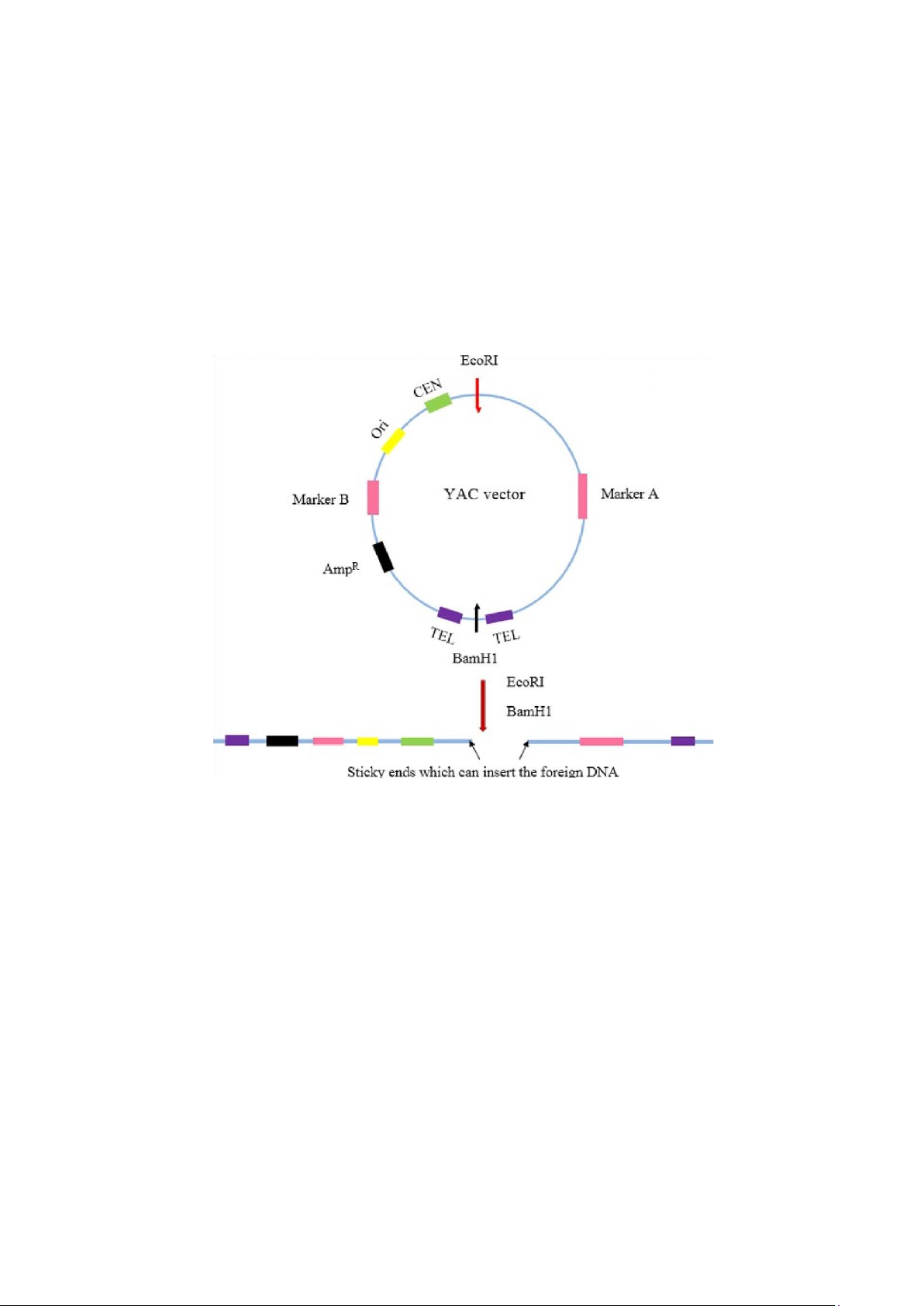
- Vector chuyển là các nhiễm sắc thể nhân tạo của nấm men YAC (Yeast
Artificial Chromosomes)
Do nhu cầu tạo dòng với những trình tự ADN ngày càng lớn, các nhà nghiên cứu
tìm tòi phát triển những vector ngày càng mới. Ngày nay người ta đã tạo ra YAC
cho phép tạo dòng với những đoạn ADN dài 150÷1.000kb, trung bình là 350kb.
Bằng nghiên cứu cho thấy ở nấm men (Saccharomyces Cerevisiae), nhiễm sắc thể
muốn nhân đôi và phân ly tốt cần ba trình tự.
YAC làm vector chuyển những đoạn gen có kích thước lớn
- 2 TEL (telomere): Trình tự đầu cuối của NST
- CEN (centrmere - tâm động): Trình tự trung tâm của NST, đảm bảo sự chia đôivà
đi về 2 cực của tế bào
- ARS (autonomously replicating sequence): Trình tự sao chép tự chủ tương tựnhư ori ở plasmid
Dựa vào đó người ta đã cấu tạo NST nhân tạo có đủ ba trình tự nói trên với cấu
tạo gồm 2 cánh tay, giữa 2 cánh tay người ta có thể cài đặt một đoạn ADN cần tạo
dòng với kích thước khoảng 150 đến 1.000kb. Trên mỗi cánh tay gồm các gen
đánh dấu di truyền để chọn lọc các tế bào nấm men có chứa YAC và các chuỗi tận
cùng có chức năng telomer đoạn cuối của NST. Một trong hai cánh tay mang một
mảnh ADN hoạt động như một tâm động và một nguồn tái bản ori. Việc cài ADN
lạ vào gen mã hóa chất ức chế tRNA vận chuyển tyrsine sẽ làm biến đổi màu sắc
khuẩn lạc tế bào gốc có mang gen hổ phách (khuẩn lạc từ trắng sang đỏ) khi có
mặt của ADN lạ. Đây là dấu hiệu về sự hiện diện của YAC tái tổ hợp trong tế bào nấm men.
Các nhiễm sắc thể này được đưa vào tế bào chủ (nấm men), nó được nhân lên như
NST khác ở trong tế bào nấm men và ta có được những gen mong muốn.
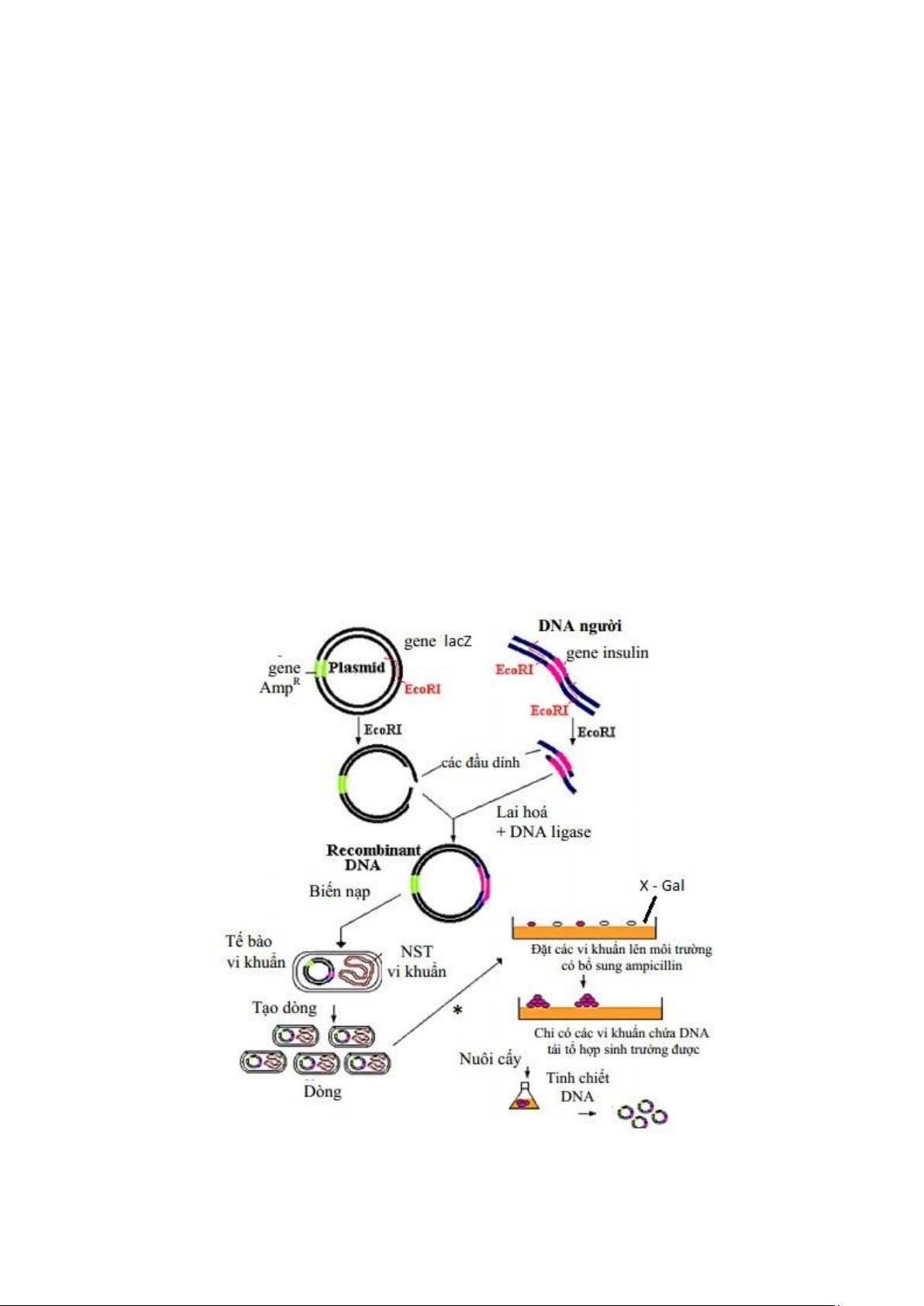
1.3. Kỹ thuật nhân dòng
Kỹ thuật tách dòng bao gồm việc phải cài một mảnh (chuỗi) ADN lạ vào
một vector (plasmide hoặc phage λ) bằng phương pháp hóa sinh. Sau đó, đưa phân
tử lai này vào tế bào chủ đã chọn lựa bằng phương pháp biến nạp hoặc tải nạp.
Trường hợp muốn tạo dòng tổng hợp enzyme thì mảnh ADN định cài phải mã hóa
cho gen cấu trúc của một enzyme nào đó.
Các bước của kỹ thuật nhân dòng
Mời các bạn cùng theo dõi những bước cơ bản trong quá trình nhân dòng qua link video sau:
https://www.youtube.com/watch?
v=DiUH4nCUSV0&list=PLjFxRogufQiNh8eJtI5k0jpZxC4UFxxZC&index=3&t=25s
1.3.1. Tạo ADN tái tổ hợp (Vector tái tổ hợp)
Việc tạo ADN tái tổ hợp bằng cách ghép ADN lạ vào vector đã được cắt cùng một
enzyme cắt hạn chế loại II, khi đó, chúng sẽ ghép đôi những đầu dính lại với nhau
nhờ bắt cặp bổ sung. Một phản ứng ghép nối xảy ra với sự có mặt của enzyme
ADN ligase của E. Coli hoặc phage T4 để hoàn chỉnh phân tử lai. Tách lập các
ADN lạ cần tạo dòng: Chọn và cắt ADN lạ của tế bào cho bằng một enzyme cắt
hạn chế (RE). Phân lập đoạn ADN (gen quí) phù hợp với vector và mục đích cần
tạo dòng. Trong trường hợp đặc biệt, để tạo dòng một gen chưa biết, người nghiên
cứu có thể tổng hợp hóa học đoạn ADN cần tạo dòng khi dự đoán cấu trúc protein
do gen chỉ huy tổng hợp hoặc tổng hợp cADN từ mRNA. Chọn và xử lí vector:
Chọn vector cần phải chú ý những yêu cầu sau: độ lớn của gen lạ (đoạn cài), loại
tế bào chủ tiếp nhận vector và phương pháp ứng dụng. Cắt vector bằng enzyme
cắt hạn chế cùng loại với enzyme đã cắt ADN nói trên (để tạo những vết cắt giống
nhau, thuận tiện cho việc nối ghép sau này). Khử nhóm phosphat bằng enzyme
alkanline phosphotase để tránh hai đầu vector đóng kín trở lại.
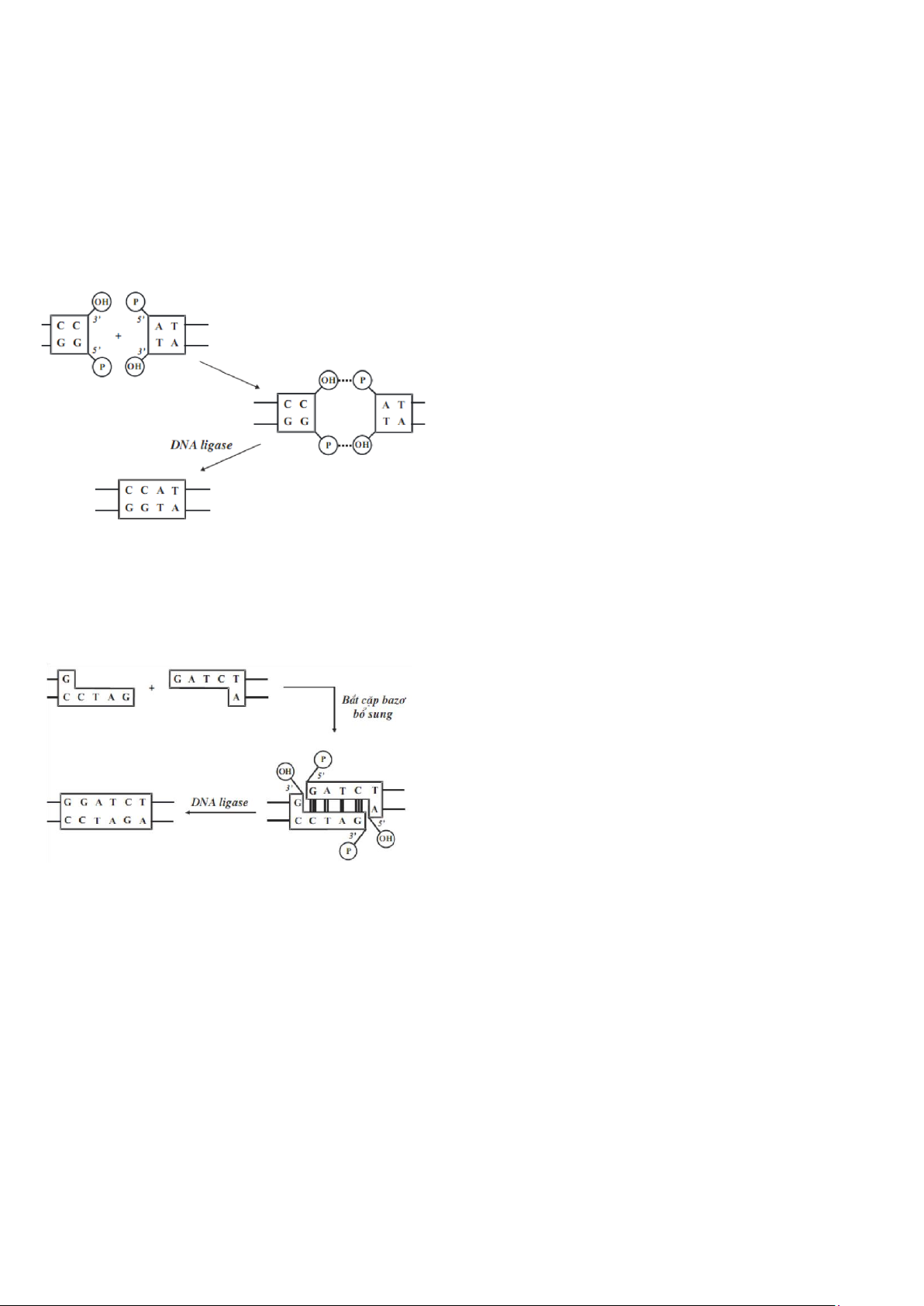
Công đoạn nối với sự có mặt của ADN ligase: Bước tiếp theo của kĩ thuật di truyền là nối các
đoạn ADN vào vector chuyển gen để tạo plasmid có mang ADN lạ. Phản ứng nối được thực
hiện nhờ enzyme ADN ligase. ADN ligase là một enzyme xúc tác các phản ứng nối hai mảnh
ADN bằng cách tạo cầu nối phosphodiester giữa đầu 5’(P) và đầu 3’(OH) của hai nucleotide
đứng cạnh nhau (Hình 6-2). Trong sinh học phân tử, người ta coi ADN ligase như một chất keo
phân tử để kết dính các mẫu ADN lại với nhau.
Nối đầu bằng
Khi hai đoạn ADN đầu bằng đứng cạnh nhau. Dưới tác dụng của enzyme ligase,
liên kết phosphodiester giữa đầu 5’(P) và đầu 3’(OH) được hình thành và hai
nucleotide được nối lại với nhau. Khả năng nối hai đoạn ADN đầu bằng rất thấp.
Để tăng hiệu suất phản ứng, thường người ta phải tăng nồng độ của các đoạn ADN.
Nối đầu lệch
Đầu tiên, hai mảnh ADN đầu lệch do có những bazơ bổ sung nên chúng tiến gần
lại nhau để tạo liên kết hydro giữa các bazơ nitơ bổ sung. Sau đó, hai nucleotide
của hai đầu nối tạo liên kết este giữa OH(3’) và P(5’) dưới tác dụng của enzyme ADN ligase.
ADN và các phản ứng nối
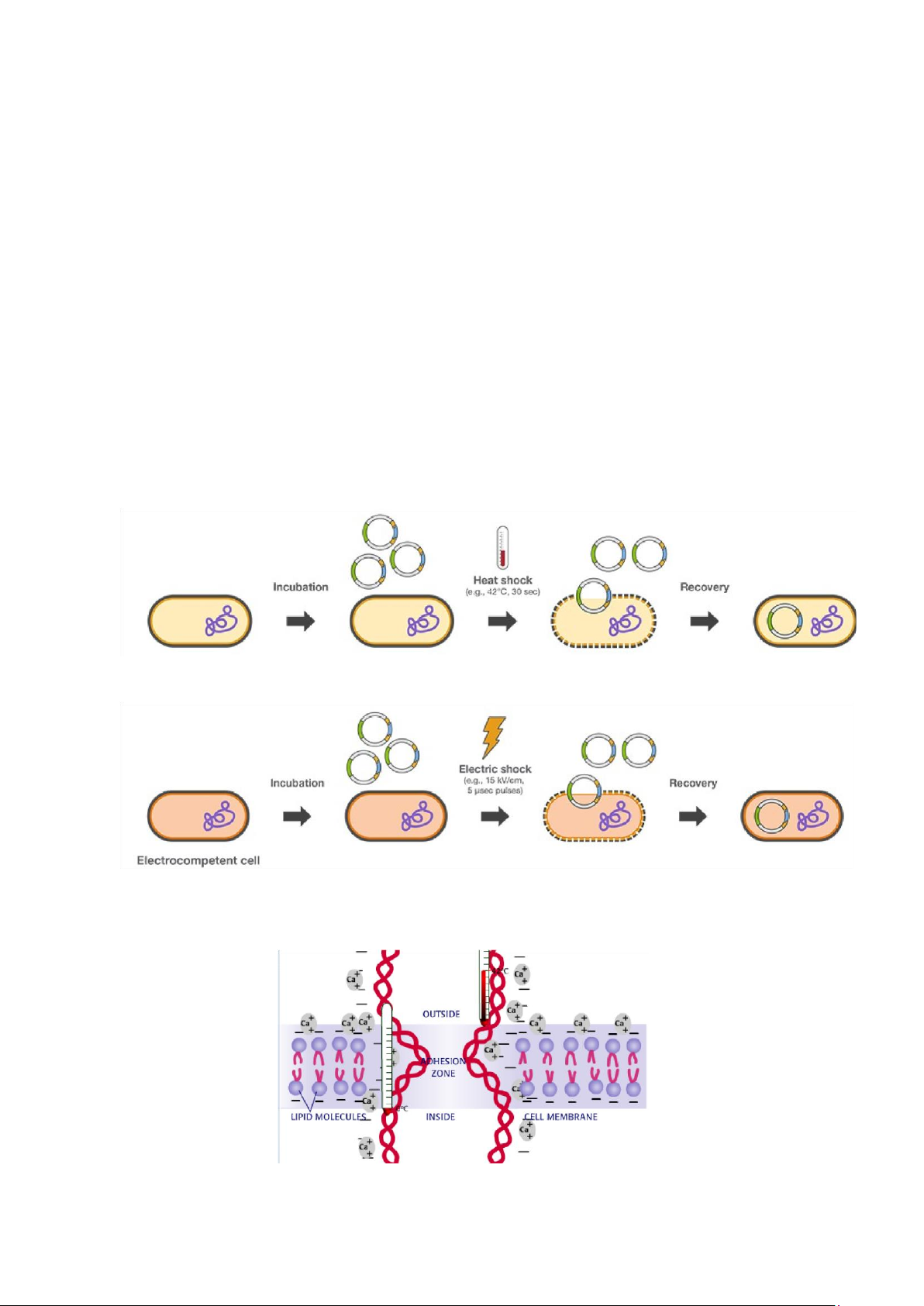
1.3.2. Chuyển ADN tái tổ hợp vào tế bào chủ bằng phương pháp biến nạp hoặc tải nạp
Công đoạn này nhằm mục đích sử dụng bộ máy của tế bào chủ để sao chép
vector tái tổ hợp thành một số lượng lớn bản sao. Việc chuyển ADN tái tổ hợp vào
tế bào vi khuẩn tức là làm cho vi khuẩn trở thành khả biến, nghĩa là có khả năng
thấm vector tái tổ hợp. Sự thấm này có thể xảy ra một cách tự nhiên hoặc được
cảm ứng. Tuy nhiên, nó sẽ phụ thuộc vào loại plasmid sử dụng làm vector và phụ
thuộc vào sự định vị của vùng cài lắp chứa bên trong vector mà người nghiên cứu
sẽ chọn phương pháp biến nạp hoặc tải nạp.
Biến nạp là hiện tượng chuyển vật chất di truyền trực tiếp từ tế bào thể cho
D (Doner) sang tế bào thể nhận R (Reception), không cần sự tiếp xúc giữa hai tế
bào hoặc nhân tố trung gian là phage hoặc virus. Biến nạp được thực hiện với
vector chuyển gen là plasmid. Có nhiều phương pháp biến nạp như hóa biến nạp,
điện biến nạp, biến nạp tế bào trần, phương pháp bắn gen và phương pháp vi tiêm.
Tải nạp là hiện tượng chuyển vật chất di truyền trực tiếp từ tế bào thể cho
D (Doner) sang tế bào thể nhận R (Reception) qua nhân tố trung gian là virus. Tải
nạp được thực hiện với vector chuyển gen có nguồn gốc là virus như phage, cosmid, ...
Chuyển plasmid vào tế bào bằng cách xử lý CaCl2 ở nhiệt độ 42oC trong 30 giây
Chuyển plasmid vào tế bào bằng cách dùng xung điện ở 15kV/cm trong 5.10-3 giây
Plasmid được đưa vào tế bào nhận bằng cách xử lý hóa học
Tài liệu liên quan:
-

Bài giảng sinh học di truyền | Trường Đại học Đồng Tháp
11 6 -

Bài tập Di truyền xác suất và di truyền người môn Sinh học di truyền | Đại học Đồng Tháp
50 25 -

Đề cương ôn thi Sinh học phân tử và di truyền phân tử môn Sinh học di truyền | Đại học Đồng Tháp
66 33 -

Trắc nghiệm môn Sinh học di truyền | Trường Đại học Đồng Tháp
380 190