Lý thuyết Enzyme môn Sinh học tế bào | Đại học Bách Khoa Hà Nội
Lý thuyết Enzyme môn Sinh học tế bào | Đại học Bách Khoa Hà Nội. Tài liệu được sưu tầm giúp bạn tham khảo, ôn tập và đạt kết quả cao. Mời bạn đọc đón xem.
Môn: Sinh học tế bào 19 tài liệu
Trường: Đại học Bách Khoa Hà Nội 5.8 K tài liệu
Tác giả:







Preview text:
Table of Contents
I. Định nghĩa..................................................................................................................... 1 II.
Cơ chế hoạt động của PGPR. ..................................................................................... 1 1.
Cơ chế hoạt động trực tiếp:...................................................................................................2 1.1.
Cố định nito .......................................................................................................................................... 3 1.2.
Hoà tan phosphate................................................................................................................................ 3 1.3.
Sản xuất phytohormone ....................................................................................................................... 4 1.4.
Sản xuất siderophore ........................................................................................................................... 6 2.
Cơ chế hoạt động gián tiếp: ..................................................................................................7 2.1.
Sản xuất hydro xyanua (HCN)............................................................................................................ 8 2.2.
Sản xuất enzyme thủy phân bởi PGPR .............................................................................................. 8 2.3.
Sản xuất kháng sinh bởi PGPR........................................................................................................... 9 2.4.
Kháng hệ thống cảm ứng (ISR) .......................................................................................................... 9 III.
Cách thu nhận enzyme. ............................................................................................ 10 IV.
Ứng dụng của enzyme. ............................................................................................. 10
V. Tài liệu tham khảo. ...................................................................................................... 10 I. Định nghĩa.
II. Cơ chế hoạt động của PGPR.
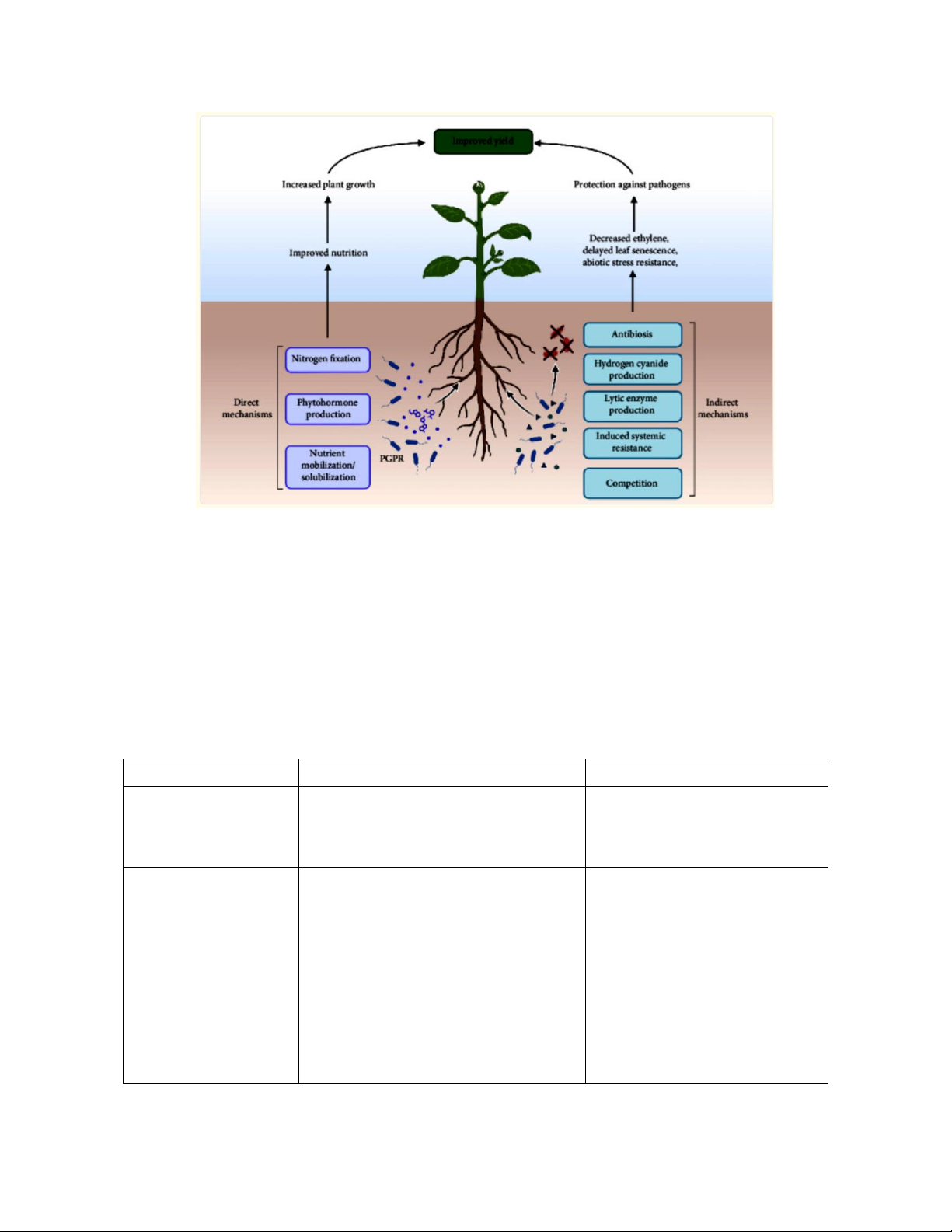
PGPR luôn hoạt động thông qua hai cơ chế hoạt động: gián tiếp và trực tiếp. Cơ
Hình 1: Sơ đồ mô tả cơ chế hoạt động của PGPR
1. Cơ chế hoạt động trực tiếp:
Cơ chế tác động trực tiếp ảnh hưởng đến sự cân bằng tăng trưởng của cây trồng
thông qua các chất điều hoà sinh trưởng. PGPR sản xuất các chất điều hoà này,
sau đó hệ thống thực vật sẽ tích hợp chúng. Không những thế, PGPR cung cấp
các yếu tố thiết yếu cho cây như phytohormone, cố định đạm và hòa tan khoáng
chất, giúp cây hấp thu dinh dưỡng dễ dàng và hiệu quả hơn. PGPR Định nghĩa Cơ chế hoạt động
Chất kích thích Đây là một nhóm vi khuẩn ở Sản xuất phytohormone sinh trưởng
vùng rế có khả năng sản xuất các phytohormone.
Phân bón sinh Đây là các vi sinh vật sống, Hoà tan các phosphat vô học
khi được bón cho cây sẽ xâm cơ, cố định nito
chiếm môi trường vi mô của
rễ, từ đó kích thuchs sự sinh
trưởng của cây thông qua
việc tăng khả năng hấp thụ
các chất dinh dưỡng thiết yếu cho cây.
Thuốc trừ sâu Đây là những vi sinh vật giúp Sản xuất enzyme phân sinh học
thúc đẩy sự phát triển của cây giải; sản xuất
bằng cách đối kháng với các siderphore, kháng sinh,
tác nhân gây bệnh có hại cho hỷo xyanua cây.
Việc thúc đẩy sự phát triển của cây trồng khi không có tác nhân gây bệnh được
xếp vào loại tác động trực tiếp của PGPR. Điều này bao gồm việc tạo ra điều
kiện thuận lợi cho việc hấp thụ chất dinh dưỡng và sản xuất phytohormone
thực vật, ảnh hưởng đến cả vùng rễ và vùng lá. 1.1. Cố định nito
Các chủng PGPR như Bacillus azotofixans có khả năng chuyển đổi đạm khí
quyển thành dạng amoniac, cung cấp nguồn đạm dễ tiêu cho cây.
Các vi khuẩn vùng rễ có khả năng cố định nitơ khí quyển được chia thành hai
nhóm chính: vi khuẩn cộng sinh và vi khuẩn cố định đạm sống tự do.
1.1.1. Cố định nito cộng sinh:
Vi khuẩn cộng sinh, điển hình là nhóm liên kết với cây họ đậu, xâm nhập vào
rễ và hình thành nốt sần để cố định nitơ (Rhizobium, Bradyrhizobium)
1.1.2. Cố định nito tự do:
Vi khuẩn cố định đạm sống tự do tạo mối liên hệ không đặc hiệu và lỏng lẻo
với rễ cây, nhưng vẫn góp phần cung cấp nitơ cho cây trồng. Nhiều chủng
PGPR thuộc các chi Azospirillum và Azotobacter đã được ghi nhận rộng rãi
nhờ khả năng cố định đạm cả trong điều kiện phòng thí nghiệm và ngoài môi trường.
- Không cần cộng sinh (Azotobacter, Azospirillum, Pseudomonas)
- Chuyển đổi N₂ trong không khí thành NH₄⁺ có thể sử dụng được
- Đóng góp 65% lượng nitơ trong nông nghiệp
1.2. Hoà tan phosphate
Phốt pho là nguyên tố dinh dưỡng đa lượng thiết yếu cho thực vật nhưng phần
lớn tồn tại trong đất ở dạng không hòa tan, khiến cây khó hấp thụ. Thực vật chỉ
sử dụng được phốt pho ở dạng phosphat đơn (H₂PO₄⁻) và phosphat kép
(HPO₄²⁻). Trong đất, các vi khuẩn thúc đẩy tăng trưởng thực vật (PGPR) có
khả năng chuyển hóa các phosphat không tan như phosphat nhôm, phosphat
canxi, đá phosphat và phosphat hữu cơ phức tạp thành dạng vô cơ hòa tan, giúp
cây dễ dàng hấp thụ hơn.
PGPR hòa tan phosphat chủ yếu thông qua giải phóng axit hữu cơ trong quá
trình chuyển hóa carbohydrate, làm giảm pH cục bộ và phá vỡ liên kết
phosphat–cation. Nhóm vi khuẩn này chiếm khoảng 20–40% quần thể vi sinh
vật trong đất, tập trung nhiều nhất ở vùng rễ và thể hiện hoạt tính trao đổi chất
cao so với vi khuẩn ngoài vùng rễ.
Khả năng đồng hóa phốt pho của cây bị hạn chế do sự kết tủa phosphat với
Fe³⁺, Al³⁺ và Ca²⁺: trong đất chua, phosphat chủ yếu tồn tại dưới dạng muối sắt
và nhôm, còn trong đất đá vôi là canxi phosphat. Phốt phát hữu cơ chiếm tỷ lệ
lớn trong đất giàu chất hữu cơ, trong đó phytate chiếm tới ~80%. Mặc dù một
số vi sinh vật sản xuất enzyme phytase có thể thủy phân phytate, hiệu quả của
enzyme này thường bị hạn chế do tạo phức với các ion kim loại như Fe, Al và Ca.
1.3. Sản xuất phytohormone
Phytohormone (auxin, cytokinin, gibberellin, ethylene, axit abscisic,
brassinosteroid và jasmonate) là các hormone thực vật điều hòa nhiều quá trình
sinh trưởng và phát triển. Vi sinh vật đất, đặc biệt là vi khuẩn vùng rễ (PGPR),
có khả năng sản xuất phytohormone hiệu quả và cây trồng có thể dễ dàng đồng
hóa các hormone này, từ đó thúc đẩy phân chia tế bào, kéo dài rễ và hình thành
hệ rễ ở cả cây họ đậu và không họ đậu.
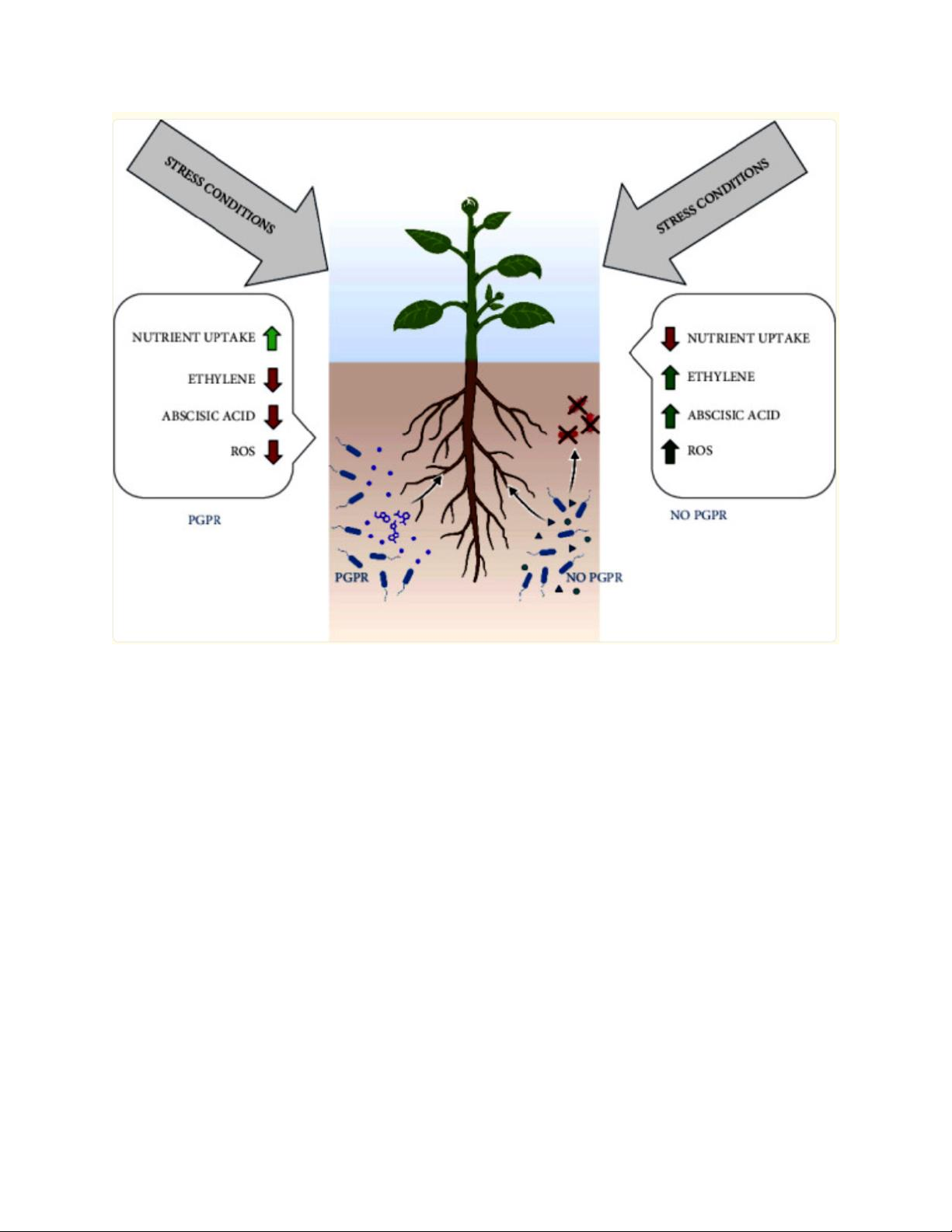
Hình 2: Cơ chế cải thiện khả năng chịu đựng của cây trồng đối với các tác
động bất lợi từ môi trường. Mũi tên màu xanh lá cây biểu thị hiệu ứng kích
hoạt, trong khi mũi tên màu đỏ biểu thị hiệu ứng giảm thiểu.
Sản xuất hormone thực vật: 1.3.1. Auxin (IAA):
Auxin, chủ yếu là axit indole-3-acetic (IAA), là phytohormone được hơn 80%
PGPR sản xuất. IAA đóng vai trò trung tâm trong kéo dài tế bào, phân chia tế
bào và phát triển hệ rễ. Auxin do PGPR tạo ra kết hợp với auxin nội sinh của
cây tạo hiệu ứng cộng hưởng, giúp hệ rễ phát triển mạnh, tăng diện tích tiếp
xúc với đất và cải thiện khả năng hấp thụ dinh dưỡng. IAA được tổng hợp qua
các con đường phụ thuộc hoặc không phụ thuộc L-tryptophan, trong đó con
đường phụ thuộc L-tryptophan phổ biến hơn. 1.3.2. Cytokinin:
Cytokinin do PGPR sản xuất kích thích phân chia tế bào, hình thành lông rễ và
chồi, đồng thời ức chế sự kéo dài rễ. Cytokinin chủ yếu tồn tại dưới dạng zeatin
hoặc kinetin và được tổng hợp qua con đường AMP hoặc tRNA. Nhiều chủng
PGPR như Bacillus megaterium và Bradyrhizobium đã được chứng minh có
khả năng sản xuất zeatin. 1.3.3. Gibberellin:
Gibberellin (GA) là nhóm phytohormone điều hòa kéo dài thân, nảy mầm, ra
hoa và đậu quả. Một số PGPR có thể sản xuất các GA sinh học quan trọng như
GA₁, GA₃, GA₄ và GA₂₀. GA do PGPR tạo ra có thể vận chuyển từ rễ lên phần
trên mặt đất, thúc đẩy sinh trưởng, đặc biệt khi kết hợp với auxin. Việc sản
xuất GA đã được xác nhận bằng các phương pháp đáng tin cậy như GC–MS ở một số loài vi khuẩn.
1.4. Sản xuất siderophore
PGPR tạo ra siderophore, một hợp chất có khả năng liên kết và vận chuyển sắt,
giúp cây hấp thu nguyên tố vi lượng quan trọng này một cách hiệu quả.
Sắt là chất dinh dưỡng thiết yếu cho thực vật, tham gia vào các quá trình sinh
lý quan trọng như cố định nitơ, hô hấp và quang hợp. Mặc dù sắt rất phổ biến
trong đất, nhưng dạng oxy hóa chủ yếu của nó dễ tạo thành oxit và hydroxit
không tan, khiến thực vật và vi sinh vật khó hấp thụ. Để khắc phục điều này,
thực vật đã phát triển hai cơ chế hấp thụ sắt: (i)
Tiết axit hữu cơ tạo phức hòa tan với sắt, sau đó khử và hấp thụ tại rễ.
(ii) Hấp thụ trực tiếp các phức sắt không tan và khử chúng trong hệ thống thực vật.
Nhiều vi khuẩn kích thích sinh trưởng thực vật (PGPR) sản xuất siderophore,
các hợp chất có khả năng tạo phức mạnh với sắt trong vùng rễ, giúp tăng tính
di động và khả năng hấp thụ sắt của cây. Siderophore có trọng lượng phân tử
thấp (<1 kDa), liên kết thuận nghịch với sắt và thường thuộc các nhóm
catechol, carboxylate và hydroxamate. Các chủng Pseudomonas tiết
siderophore như pyoverdine và pyochelin, giúp chúng chiếm ưu thế cạnh tranh
trong vùng rễ nhờ đặc tính kháng sinh.
Vi khuẩn PGPR sản xuất siderophore không chỉ tăng cường hấp thu sắt và thúc
đẩy sinh trưởng cây, mà còn ức chế nấm gây bệnh bằng cách làm giảm nguồn
sắt sẵn có, do nhiều tác nhân gây bệnh không thể sử dụng phức hợp sắt– siderophore. Sản xuất siderophore:
Siderophore là phân tử có khả năng kết hợp với sắt (Fe³⁺)
Tăng khả năng hấp thụ sắt cho thực vật
Cạnh tranh với vi sinh vật gây bệnh về nguồn sắt
2. Cơ chế hoạt động gián tiếp:
Cơ chế hoạt động gián tiếp tận dụng cơ chế phòng vệ tự nhiên của cây trồng
thông qua quá trình truyền tín hiệu vi sinh. Bên cạnh đất, thông qua cơ chế gián
tiếp, PGPR bảo vệ cây trồng chống lại căng thẳng môi trường.
PGPR bảo vệ cây bằng cách ức chế các vi sinh vật gây bệnh và kích thích hệ
miễn dịch tự nhiên của cây (Induced Systemic Resistance - ISR), giúp cây tăng
cường khả năng chống chịu bệnh tật.
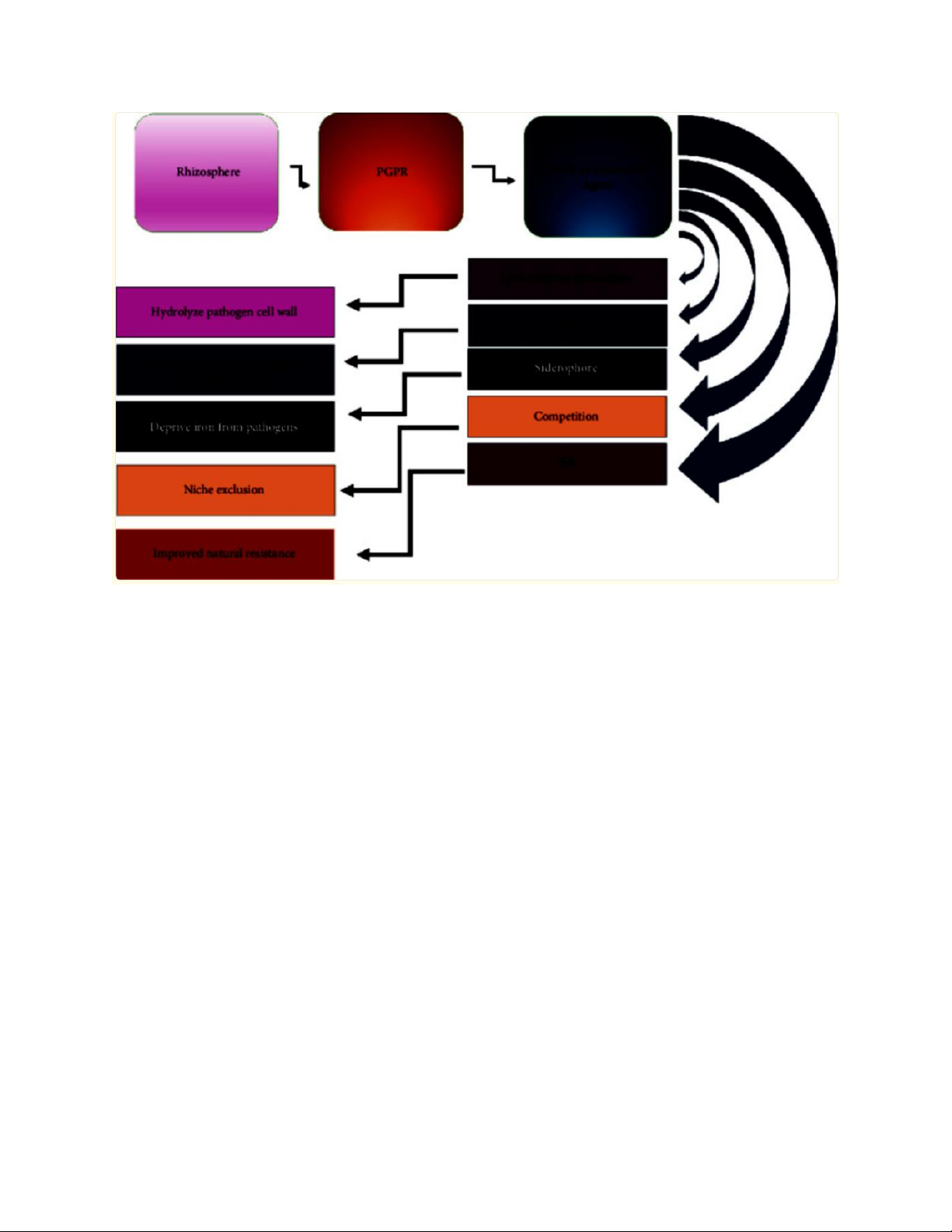
Vi khuẩn vùng rễ thúc đẩy sinh trưởng cây trồng gián tiếp bằng cách hoạt động
như tác nhân kiểm soát sinh học, thông qua cạnh tranh dinh dưỡng, loại
trừ vị trí sinh thái, cảm ứng kháng bệnh toàn thân và sản xuất các chất
chuyển hóa kháng nấm, kháng khuẩn.
Hình 3: Cơ chế hoạt động của PGPR như một tác nhân kiểm soát sinh học.
2.1. Sản xuất hydro xyanua (HCN)
PGPR có khả năng sản xuất HCN thông qua con đường sinh tổng hợp de novo,
đóng vai trò như một chất kháng khuẩn mạnh. HCN ức chế cytochrome oxidase
trong hô hấp tế bào, làm gián đoạn chuỗi vận chuyển điện tử và dẫn đến tế bào
mầm bệnh bị chết, qua đó tăng hiệu quả kiểm soát sinh học. Nhiều chủng
Pseudomonas và Bacillus là các nhà sản xuất HCN điển hình; ngoài ra, khả
năng này cũng được ghi nhận ở Burkholderia, Rhizobia và Chromobacterium,
cho thấy tính đa dạng của PGPR. Đặc biệt, Pseudomonas huỳnh quang sản xuất
HCN hiệu quả, góp phần ức chế mạnh các bệnh do Fusarium trên cà chua.
2.2. Sản xuất enzyme thủy phân bởi PGPR
PGPR kiểm soát sinh học mầm bệnh bằng cách sản xuất các enzyme phân hủy
thành tế bào như glucanase, protease, chitinase và cellulase, từ đó phá vỡ cấu
trúc thành tế bào của nấm gây bệnh. Chitinase đặc biệt quan trọng vì phân giải
chitin, thành phần thiết yếu của thành tế bào nấm. Thành tế bào của Fusarium
oxysporum có thể bị phân hủy bởi β-1,4-glucanase do các chủng Streptomyces
và Paenibacillus sản xuất. Nhiều vi khuẩn như Serratia, Pseudomonas,
Enterobacter và Bacillus thể hiện hoạt tính phân giải chitin mạnh. Đáng chú ý,
Serratia marcescens tạo chitinase có hiệu quả kháng nấm cao đối với F.
oxysporum và Rhizoctonia solani, gây biến dạng sợi nấm (đứt gãy, cuộn,
phồng), minh chứng rõ ràng cho cơ chế kiểm soát sinh học.
2.3. Sản xuất kháng sinh bởi PGPR
Việc sử dụng thiên địch vi sinh (PGPR) là giải pháp thay thế tiềm năng cho
thuốc trừ sâu hóa học. Nhiều chủng Bacillus và Pseudomonas kiểm soát mầm
bệnh hiệu quả thông qua tiết kháng sinh ngoại bào, có hoạt tính mạnh ngay cả
ở nồng độ thấp. Bacillus sản xuất nhiều hợp chất kháng khuẩn, kháng nấm
quan trọng như sublancin, subtilosin A, TasA, subtilin và các lipopeptide thuộc
họ iturin, fengycin, surfactin.
Trong khi đó, Pseudomonas aeruginosa và P. fluorescens tạo ra nhiều chất
chuyển hóa thứ cấp có hoạt tính sinh học rộng, không chỉ kháng vi khuẩn và
nấm mà còn kháng virus, chống oxy hóa, gây độc tế bào và ức chế sinh vật gây
hại. Các hợp chất tiêu biểu gồm phenazine, DAPG, pyoluteorin, pyrrolnitrin,
oomycin A, ecomycins và cepafungins, cho thấy tiềm năng lớn của PGPR
trong kiểm soát sinh học và ứng dụng nông nghiệp bền vững.
2.4. Kháng hệ thống cảm ứng (ISR)
PGPR bảo vệ thực vật gián tiếp thông qua kháng bệnh toàn thân cảm ứng (ISR),
trong đó vi khuẩn vùng rễ kích hoạt hệ thống phòng vệ nội tại của cây dù không
tiếp xúc trực tiếp với mầm bệnh. ISR có tính không đặc hiệu, giúp cây kháng
được nhiều loại tác nhân gây bệnh khác nhau, nên rất có giá trị trong điều kiện
tự nhiên và sản xuất nông nghiệp.
Một số chủng Pseudomonas đã được chứng minh kích hoạt ISR ở lúa mì và củ
cải, với tác nhân cảm ứng là chuỗi O-antigen của lipopolysaccharide màng
ngoài và các siderophore như pseudobactin và pseudomanine. Các siderophore
này có thể kích thích sinh tổng hợp axit salicylic, từ đó khởi phát ISR. Việc
cấy các PGPR cảm ứng ISR vào rễ tạo ra đáp ứng phòng vệ lan tỏa toàn cây,
giúp tăng mạnh khả năng kháng bệnh.
III. Cách thu nhận enzyme.
IV. Ứng dụng của enzyme. V.
Tài liệu tham khảo.
[1] BioSolutions for Green Agriculture: Unveiling the Diverse Roles of Plant
Growth-Promoting Rhizobacteria, 2024.
Document Outline
- I. Định nghĩa.
- II. Cơ chế hoạt động của PGPR.
- 1. Cơ chế hoạt động trực tiếp:
- 1.1. Cố định nito
- 1.2. Hoà tan phosphate
- 1.3. Sản xuất phytohormone
- 1.4. Sản xuất siderophore
- 2. Cơ chế hoạt động gián tiếp:
- 2.1. Sản xuất hydro xyanua (HCN)
- 2.2. Sản xuất enzyme thủy phân bởi PGPR
- 2.3. Sản xuất kháng sinh bởi PGPR
- 2.4. Kháng hệ thống cảm ứng (ISR)
- 1. Cơ chế hoạt động trực tiếp:
- III. Cách thu nhận enzyme.
- IV. Ứng dụng của enzyme.
- V. Tài liệu tham khảo.
Tài liệu liên quan:
-

bài giảng giải tích 1 trường bác khoa
18 9 -

N Ahmad - Clinical and etiological profile of unprovoked thrombosis in young patients admitted at a
17 9 -

Bài giảng về Sự sinh sản của tế bào Eukaryote môn Sinh học tế bào | Đại học Bách Khoa Hà Nội
37 19 -

The cell division and cell death môn Sinh học tế bào | Đại học Bách Khoa Hà Nội
49 25 -

Bài giảng Tổng quan về tế bào môn Sinh học tế bào | Đại học Bách Khoa Hà Nội
41 21